Le Passage Cénomanien
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

December 2020 Contract Pipeline
OFFICIAL USE No Country DTM Project title and Portfolio Contract title Type of contract Procurement method Year Number 1 2021 Albania 48466 Albanian Railways SupervisionRehabilitation Contract of Tirana-Durres for Rehabilitation line and ofconstruction the Durres of- Tirana a new Railwaylink to TIA Line and construction of a New Railway Line to Tirana International Works Open 2 Albania 48466 Albanian Railways Airport Consultancy Competitive Selection 2021 3 Albania 48466 Albanian Railways Asset Management Plan and Track Access Charges Consultancy Competitive Selection 2021 4 Albania 49351 Albania Infrastructure and tourism enabling Albania: Tourism-led Model For Local Economic Development Consultancy Competitive Selection 2021 5 Albania 49351 Albania Infrastructure and tourism enabling Infrastructure and Tourism Enabling Programme: Gender and Economic Inclusion Programme Manager Consultancy Competitive Selection 2021 6 Albania 50123 Regional and Local Roads Connectivity Rehabilitation of Vlore - Orikum Road (10.6 km) Works Open 2022 7 Albania 50123 Regional and Local Roads Connectivity Upgrade of Zgosth - Ura e Cerenecit road Section (47.1km) Works Open 2022 8 Albania 50123 Regional and Local Roads Connectivity Works supervision Consultancy Competitive Selection 2021 9 Albania 50123 Regional and Local Roads Connectivity PIU support Consultancy Competitive Selection 2021 10 Albania 51908 Kesh Floating PV Project Design, build and operation of the floating photovoltaic plant located on Vau i Dejës HPP Lake Works Open 2021 11 Albania 51908 -

S.No Governorate Cities 1 L'ariana Ariana 2 L'ariana Ettadhamen-Mnihla 3 L'ariana Kalâat El-Andalous 4 L'ariana Raoued 5 L'aria
S.No Governorate Cities 1 l'Ariana Ariana 2 l'Ariana Ettadhamen-Mnihla 3 l'Ariana Kalâat el-Andalous 4 l'Ariana Raoued 5 l'Ariana Sidi Thabet 6 l'Ariana La Soukra 7 Béja Béja 8 Béja El Maâgoula 9 Béja Goubellat 10 Béja Medjez el-Bab 11 Béja Nefza 12 Béja Téboursouk 13 Béja Testour 14 Béja Zahret Mediou 15 Ben Arous Ben Arous 16 Ben Arous Bou Mhel el-Bassatine 17 Ben Arous El Mourouj 18 Ben Arous Ezzahra 19 Ben Arous Hammam Chott 20 Ben Arous Hammam Lif 21 Ben Arous Khalidia 22 Ben Arous Mégrine 23 Ben Arous Mohamedia-Fouchana 24 Ben Arous Mornag 25 Ben Arous Radès 26 Bizerte Aousja 27 Bizerte Bizerte 28 Bizerte El Alia 29 Bizerte Ghar El Melh 30 Bizerte Mateur 31 Bizerte Menzel Bourguiba 32 Bizerte Menzel Jemil 33 Bizerte Menzel Abderrahmane 34 Bizerte Metline 35 Bizerte Raf Raf 36 Bizerte Ras Jebel 37 Bizerte Sejenane 38 Bizerte Tinja 39 Bizerte Saounin 40 Bizerte Cap Zebib 41 Bizerte Beni Ata 42 Gabès Chenini Nahal 43 Gabès El Hamma 44 Gabès Gabès 45 Gabès Ghannouch 46 Gabès Mareth www.downloadexcelfiles.com 47 Gabès Matmata 48 Gabès Métouia 49 Gabès Nouvelle Matmata 50 Gabès Oudhref 51 Gabès Zarat 52 Gafsa El Guettar 53 Gafsa El Ksar 54 Gafsa Gafsa 55 Gafsa Mdhila 56 Gafsa Métlaoui 57 Gafsa Moularès 58 Gafsa Redeyef 59 Gafsa Sened 60 Jendouba Aïn Draham 61 Jendouba Beni M'Tir 62 Jendouba Bou Salem 63 Jendouba Fernana 64 Jendouba Ghardimaou 65 Jendouba Jendouba 66 Jendouba Oued Melliz 67 Jendouba Tabarka 68 Kairouan Aïn Djeloula 69 Kairouan Alaâ 70 Kairouan Bou Hajla 71 Kairouan Chebika 72 Kairouan Echrarda 73 Kairouan Oueslatia 74 Kairouan -

Entreprise Code Sec Ville Siege Adresse Tel Fax
ENTREPRISE CODE_SEC VILLE_SIEGE ADRESSE TEL FAX 1 BELDI IAA ARIANA Route de Mateur Km 8 71 521 000 71 520 577 2 BISCUITERIE AZAIZ IAA ARIANA 71 545 141 71 501 412 3 COOPERATIVE VITICOLE DE TUNIS IAA ARIANA Sabalet ben Ammar 71 537 120 71 535 318 4 GENERAL FOOD COMPANY IAA ARIANA Rue Metouia BORJ LOUZIR 71 691 036 70 697 104 5 GRANDE FABRIQUE DE CONFISERIE ORIENTALE - GFCO IAA ARIANA 11, Rue des Entrepreneurs Z.I Ariana Aroport 2035 Tunis-Carthage 70837411 70837833 6 HUILERIE BEN AMMAR IAA ARIANA Cebelet Ben Ammar Route de Bizerte Km 15 71 537 324 71 785 916 7 SIROCCO IAA ARIANA Djebel Ammar 71 552 365 71 552 098 8 SOCIETE AMANI IAA ARIANA Route Raoued Km 5 71 705 434 71 707 430 9 SOCIETE BGH IAA ARIANA Z.I Elalia Ben Gaied Hassine 71 321 718/70823945 70823944 10 SOCIETE CARTHAGE AGRO-ALIMENTAIRE IAA ARIANA Bourj Touil 70684001 70684002 11 SOCIETE DE SERVICES AGRICOLES ZAHRA IAA ARIANA Bouhnech - KALAAT EL ANDALUS 25 100 200 12 SOCIETE FROMAGERIE SCANDI IAA ARIANA 23 346 143/706800 70 680 009 13 SOCIETE FRUIT CENTER IAA ARIANA 35 Rue Mokhtar ATTIA 71 334 710 71 857 260 14 SOCIETE GIGA IAA ARIANA 70 308 441 71 308 476 15 SOCIETE GREEN LAND ET CIE IAA ARIANA route el battane jedaida 1124 mannoba 71798987 71784116 16 SOCIETE JASMIN EXPORT IAA ARIANA Rue Mohamed El Habib Route de Raoued Km 7 71 866 817 71 866 826 17 SOCIETE KACEM DE PATISSERIE - KAPCO IAA ARIANA 20 Rue Kalaat Ayoub Riadh El Andalous 71 821 388 71 821 466 18 SOCIETE LABIDI VIANDES IAA ARIANA Borj Touil 71 768 731 71 769 080 19 SOCIETE LE TORREFACTEUR IAA ARIANA Rue de l'argent -

MPLS VPN Service
MPLS VPN Service PCCW Global’s MPLS VPN Service provides reliable and secure access to your network from anywhere in the world. This technology-independent solution enables you to handle a multitude of tasks ranging from mission-critical Enterprise Resource Planning (ERP), Customer Relationship Management (CRM), quality videoconferencing and Voice-over-IP (VoIP) to convenient email and web-based applications while addressing traditional network problems relating to speed, scalability, Quality of Service (QoS) management and traffic engineering. MPLS VPN enables routers to tag and forward incoming packets based on their class of service specification and allows you to run voice communications, video, and IT applications separately via a single connection and create faster and smoother pathways by simplifying traffic flow. Independent of other VPNs, your network enjoys a level of security equivalent to that provided by frame relay and ATM. Network diagram Database Customer Portal 24/7 online customer portal CE Router Voice Voice Regional LAN Headquarters Headquarters Data LAN Data LAN Country A LAN Country B PE CE Customer Router Service Portal PE Router Router • Router report IPSec • Traffic report Backup • QoS report PCCW Global • Application report MPLS Core Network Internet IPSec MPLS Gateway Partner Network PE Router CE Remote Router Site Access PE Router Voice CE Voice LAN Router Branch Office CE Data Branch Router Office LAN Country D Data LAN Country C Key benefits to your business n A fully-scalable solution requiring minimal investment -

Section: ARIANA
Section: ARIANA Nom Prénom Adresse Code postal Tél ABDELMOULA AHMED 71,Avenue Habib Bourguiba 2080 ARIANA 71716297 ABDELMOUMEN EP, OUESLATI SOUMAYA Route Principale 7024 IMADA-ZOUAOUINE 72 403 525 ABDENNEBI EP, NAKOURI LILIA 14, Avenue de la Liberté C,C,Tej 1004 EL MENZAH 5 71 237 036 ALOULOU KHEDIJA Cité Commerciale Jamil 2080 ARIANA 71754731 AMARA EP, BEN RHOUMA ZOHRA 19, Rue Taieb M'Hiri 2041 CITE ETTADHAMEN 71 516 453 AMARA EP,MEDDEB CAMELIA Bezina 7012 BAZINA AMMAR EP,KRICHEN ZEINEB 44, Avenue Taieb M'Hiri 2080 ARIANA 71714659 AMRI MOHAMED NEJIB 11,Avenue Habib Bourguiba 1110 MORNAGUIA 71.540.255 ARBI ABDELAZIZ 19, Rue d'Algérie 7030 MATEUR 72485420 ARBI DALENDA 3, Rue d'Algérie 7050 MENZEL BOURGUIBA 72 460 219 AYADI MAHJOUB 15, Rue Musset-Ang rue Algérie 7050 MENZEL BOURGUIBA 72.463.768 AYADI EP, BEN HASSEN FADHILA 112, Avenue HabibBourguiba 2022 KALAAT EL ANDALOUS 71 558 423 AZAIEZ RIDHA Avenue Habib Bourguiba 1124 JEDEIDA 71539110 AZOUZ OLFA Résidence les Orangers- Av, des Orangers 2010 LA MANOUBA 71 603 755 AZOUZ EP, GHORBAL HAGER 1, Avenue de l'Environnement 2021 OUED ELLIL 71535301 AZOUZI EP, FERCHICHI RIM 57, Avenue Taieb M'Hiri 2041 CITE ETTADHAMEN 71 549 230 AZZOUZ ZOUHAIER 19, Avenue Emir Abdelkader- El Bhira 7000 BIZERTE 72 531 136 BACCOUCHE FERID 38, Avenue du 1er Mai 7000 BIZERTE 72 431 113 BAHRI RYM 61, Avenue Habib Bourguiba 7010 SEJNANE 7256114 BAKLOUTI EP, DJEMAL MERIAM Avenue 7 Novembre 7080 MENZEL JEMIL 72 490 600 BAKTACHE OTHMAN 19, Avenue Taieb M'Hiri 7000 BIZERTE 72 431 208 BANANI EP, M'ZAH AMENA 1, Rue de la -

Updated Checklist and Distribution of Large Branchiopods (Branchiopoda: Anostraca, Notostraca, Spinicaudata) in Tunisia
Biogeographia – The Journal of Integrative Biogeography 31 (2016): 27–53 Updated checklist and distribution of large branchiopods (Branchiopoda: Anostraca, Notostraca, Spinicaudata) in Tunisia FEDERICO MARRONE1,*, MICHAEL KORN2, FABIO STOCH3, LUIGI NASELLI- FLORES1, SOUAD TURKI4 1 Dept. STEBICEF, University of Palermo, via Archirafi, 18, I-90123 Palermo, Italy 2 Limnological Institute, University of Konstanz, Mainaustr. 252, D-78464 Konstanz & DNA-Laboratory, Museum of Zoology, Senckenberg Natural History Collections Dresden, Königsbrücker Landstrasse 159, D- 01109 Dresden, Germany 3 Dept. of Life, Health & Environmental Sciences, University of L’Aquila, Via Vetoio, I-67100 Coppito, L'Aquila, Italy 4 Institut National des Sciences et Technologies de la Mer, Rue du 02 mars 1934, 28, T-2025 Salammbô, Tunisia * e-mail corresponding author: [email protected] Keywords: Branchinectella media, fauna of Maghreb, freshwater crustaceans, Mediterranean temporary ponds, regional biodiversity. SUMMARY Temporary ponds are the most peculiar and representative water bodies in the arid and semi-arid regions of the world, where they often represent diversity hotspots that greatly contribute to the regional biodiversity. Being indissolubly linked to these ecosystems, the so-called “large branchiopods” are unanimously considered flagship taxa of these habitats. Nonetheless, updated and detailed information on large branchiopod faunas is still missing in many countries or regions. Based on an extensive bibliographical review and field samplings, we provide an updated and commented checklist of large branchiopods in Tunisia, one of the less investigated countries of the Maghreb as far as inland water crustaceans are concerned. We carried out a field survey from 2004 to 2012, thereby collecting 262 crustacean samples from a total of 177 temporary water bodies scattered throughout the country. -

Direction Régionale Bureau De Poste Code Postal Direction Régionale
Liste des bureaux de poste assurant la souscription à l’emprunt obligataire national (684 bureaux) Direction Code Direction Code Bureau de poste Bureau de poste Régionale postal Régionale postal Ariana Cité Ennasr Ariana 2001 Kairouan Chebika 3121 Ariana Géant 2002 Kairouan Cité Hajjem 3129 Ariana Sidi Thabet 2020 Kairouan Haffouz 3130 Ariana Kalaat El Andalous 2022 Kairouan Kairouan Sud 3131 Ariana Borj Baccouche 2027 Kairouan Sisseb 3132 Ariana Cebelet Ben Ammar 2032 Kairouan Karma 3133 Ariana Tunis Carthage 2035 Kairouan Kairouan Okba 3140 Ariana Soukra 2036 Kairouan El Ala 3150 Ariana Menzah 8 2037 Kairouan Hajeb Laayoune 3160 Ariana Cité Ettadhamen 2041 Kairouan Nasrallah 3170 Ariana Raoued 2056 Kairouan bouhajla 3180 Ariana Chorfech 2057 Kairouan Cite ennasr kairouan 3182 Ariana Riadh El Andalos 2058 Kairouan Rakada 3191 Ariana Borj Louzir 2073 Kairouan borji 3198 Ariana Ariana 2080 Kairouan Cité Ibn Jazzar 3199 Ariana Borj Touil 2081 Kef Kef 7100 Ariana Cité La Gazelle 2083 Kef Enneber 7110 Ariana Complexe technologique 2088 Kef Touiref 7112 Ariana Menzah 6 2091 Kef El Kalaa Khasba 7113 Ariana Mnihla 2094 Kef Jrissa 7114 Ariana Ettadhamen 2 2095 Kef Kef Ouest 7117 Kasserine Kasserine 1200 Kef Essakia 7120 Kasserine Tela 1210 Kef Borj Elaifa 7122 Kasserine BOUCHEBKA 1213 Kef Kalaat Snen 7130 Kasserine Majel Belabbès 1214 Kef Tejerouin 7150 Kasserine TLABET 1215 Kef Menzel Salem 7151 Kasserine Elaayoune 1216 Kef El Ksour 7160 Kasserine Foussana 1220 Kef Dahmani 7170 Kasserine HIDRA 1221 Kef Sers 7180 Kasserine Kasserine Nour 1230 -

L'étude Nationale Sur L'état Et La Sensibilité À La Désertification
Introduction La Tunisie se caractérise par l’ampleur des processus de désertification ayant pour origine les caractéristiques climatiques, édaphiques, géomorphologiques et socio-économiques. En effet, on estime que la superficie menacée est estimée à 94% du territoire du pays. Ainsi, la lutte contre ce fléau fût entamé depuis fort longtemps. Au cours des dernières décennies, de vastes programmes à divers objectifs tels que la protection des terres agricoles, la lutte contre la dégradation des sols, le développement et la promotion de l’agriculture paysanne, ont été amplifiés à travers l’ensemble du pays dans le cadre des projets de développements intégrés et qui s’inscrivent dans le cadre du plan économique et social. Ces divers programmes se caractérisent par de vastes panoplies d’approches et de paquets techniques. Des actions d’aménagement et de préservation des ressources naturelles ont permis d atténuer les effets de dégradation du milieu. Des efforts ont été déployés par l’Etat en plus d’une évolution institutionnelle et juridique en rapport avec LCD pour atteindre les objectifs tracés par les différentes stratégies. Ces réalisations s’intègrent dans le cadre du PAN/LCD telles que les travaux anti-érosifs et de protection des terres agricoles et de reboisement et gestion et de préservation des ressources pastorales. Divers programmes ont été financés par plusieurs organismes nationaux et bailleurs de fonds. Basé sur l’implication de la population, afin de les faire participer activement, les pouvoirs publics sont conscients de l’urgence de l’intervention pour la consolidation des investissements réalisés et l’extension des zones d’intervention jugées prioritaires. -
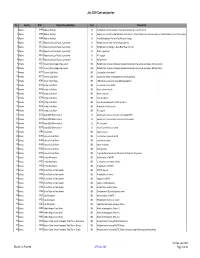
July 2020 Contract Pipeline.Xlsm
July 2020 Contract pipeline No Country DTM Project title and Portfolio Team Contract title Number 1 Albania 48466 Albanian Railways IE Rehabilitation of Tirana-Durres line and construction of a new link to TIA 2 Albania 48466 Albanian Railways IE Supervision Contract for Rehabilitation of the Durres - Tirana Railway Line and construction of a New Railway Line to Tirana Internat 3 Albania 48466 Albanian Railways IE Asset Management Plan and Track Access Charges 4 Albania 50123 Regional and Local Roads Connectivity IE Rehabilitation of Vlore - Orikum Road (10.6 km) 5 Albania 50123 Regional and Local Roads Connectivity IE Rehabilitation of Shëngjin – Baks-Rrjoll Road (15.1km) 6 Albania 50123 Regional and Local Roads Connectivity IE Works supervision 7 Albania 50123 Regional and Local Roads Connectivity IE PIU support 8 Albania 50123 Regional and Local Roads Connectivity IE Design review 9 Armenia 43253 Yerevan Water Supply Improvement IEA Rehabilitation of water distribution network and individual household connections Silikyan district 10 Armenia 43253 Yerevan Water Supply Improvement IEA Rehabilitation of water distribution network and individual household connections Malatia district 11 Armenia 46172 Yerevan Solid Waste IEA Construction of new landfill 12 Armenia 46172 Yerevan Solid Waste IEA Specialised vehicles and equipment for landfill operation 13 Armenia 46540 Gyumri Urban Roads IEA LED luminaries and other street lighting equipment 14 Armenia 46736 Kotayk Solid Waste IEA Construction of new landfill 15 Armenia 46736 Kotayk Solid -

Rapport National Du Secteur De L'eau
République Tunisienne Ministère de l’Agriculture, des Ressources Hydrauliques et de la Pêche Bureau de la Planification et des Equilibres Hydrauliques RAPPORT NATIONAL DU SECTEUR DE L’EAU Année 2018 novembre 2019 Table des matières RESUME .......................................................................................................................................... 1 Introduction .......................................................................................................................................... 7 PARTIE 1 : ORIENTATIONS POLITIQUES, CADRES REGLEMENTAIRE ET INSTITUTIONNEL I. PRINCIPAUX CONSTATS ET ORIENTATIONS, LIES AUX RESSOURCES EN EAU EN TUNISIE ........................................................................................................................................ 11 I.1. Principaux constats ....................................................................................................................... 11 I.2. Principales orientations ................................................................................................................ 13 II. LES PROGRAMMES D’APPUI INSTITUTIONNELS ET BUDGETAIRES ....................... 14 II.1. Les appuis de l’UE ........................................................................................................................ 14 II.1.1. Le programme d’appui aux politiques publiques de gestion des ressources en eau pour le développement rural et agricole PAPS-Eau .................................................................................. -

Rapport National Du Secteur De L'eau
Photo de couverture: Barrage Lakhmess République Tunisienne Ministère de l’Agriculture, des Ressources Hydrauliques et de la Pêche Bureau de la Planification et des Equilibres Hydrauliques RAPPORT NATIONAL DU SECTEUR DE L’EAU Année 2017 RAPPORT NATIONAL DU SECTEUR DE L’EAU- 2017 Table des matières RESUME .............................................................................................................................. 1 Introduction .............................................................................................................................. 9 PARTIE 1 : ORIENTATIONS POLITIQUES, CADRES REGLEMENTAIRE ET INSTITUTIONNEL I. RAPPELS DES ENJEUX ET STRATEGIES LIES AUX RESSOURCES ........................ 13 EN EAU EN TUNISIE ............................................................................................................ 13 I.1. Les enjeux lies aux ressources en eau en Tunisie .............................................................. 13 I.1.1. Mettre en œuvre la gestion de la demande .............................................................. 13 I.1.2. Assurer une gouvernance plus perfectionnée de l’eau ............................................ 13 I.1.3. Maîtriser les phénomènes extrêmes : inondations et sécheresse. ............................ 14 I.1.4. Progresser vers une gestion durable des nappes souterraines. ................................. 14 I.1.5. Améliorer la valorisation des périmètres irrigués tout en maîtrisant l’économie d’eau .......................................................................................................................................... -

Cartes Topographiques.Pdf
Les Cartes Topographiques Disponibles dans la Bibliothèque de l’INP A CA 213 Adjim: 1/100 000 Service géographique de l’armée.-Paris: S. G. A.-(Tunisie ; LXXXIV). CA 112 Agareb (Tunisie): 1/50 000 Office de la topographie et de la cartographie ; Projection Lambert.-Tunis: O. T. C.- (Tunisie: type 1982; CVI –BB -C 38). CA 140 Ain Djeloula (Tunisie): 1/ 50 000 Institut Géographique National ; Projection Lambert.- Paris: IGNF, 1957.-(Tunisie; LV- B7- C36). CA 111 Ain EL Kseiba (Tunisie): 1/50 000 Service géographique de l’armée.-Paris: S. G. A.-(Tunisie ; LX- BB - C33). CA 110 Aîne ZARETH (Tunisie): 1/100.000 Service géographique de l’armée.-Paris: S. G. A.-(Tunisie ; CXV). CA 109 Alouet Gounna (Tunisie) : 1/100.000 Service géographique de l’armée.-Paris: S. G. A.-(Tunisie ; CII). CA 108 ARIANA (Tunisie): 1/50.000 Office de la topographie et de la cartographie ; Projection Lambert.-Tunis : O. T. C.- (Tunisie ; XIII-B 1- C 36). B CA 239 Barrage Sidi Saad (Zone inondable à la côte 298): 1/10 000 Tunis : service de l’Aménagement foncier, 1978. CA 238 Barrage de Sidi Salem: 1/10.000 Tunis: service de l’Aménagement foncier, 1978. CA 107 BEJA (Tunisie): 1/50 000 Direction de la topographie et de la cartographie.-Tunis : D.T.C.-(Tunisie; XVIII). CA 106 Ben Gardane (Tunisie): 1/100 000 Service géographique national.-Paris : S.G.N.-(TUNISIE; CI). CA 105 Bir Ali Ben KHALIFA (Tunisie): 1/50 000 Institut Géographique National.- Projection Lambert.- Paris : IGNF., 1960.-(Tunisie, CV-B 1b - c 37) CA 92 Bir El GUECIRA (Tunisie): 1/100 000 Service géographique de l’armée.-Paris: S.