Birds on San Clemente Island, As Part of Our Work Toward the Recovery of the Island’S Endangered Species
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

California's Freshwater Fishes: Status and Management
California’s freshwater fishes: status and management Rebecca M. Quiñones* and Peter B. Moyle Center for Watershed Sciences, University of California at Davis, One Shields Ave, Davis, 95616, USA * correspondence to [email protected] SUMMARY Fishes in Mediterranean climates are adapted to thrive in streams with dy- namic environmental conditions such as strong seasonality in flows. Howev- er, anthropogenic threats to species viability, in combination with climate change, can alter habitats beyond native species’ environmental tolerances and may result in extirpation. Although the effects of a Mediterranean cli- mate on aquatic habitats in California have resulted in a diverse fish fauna, freshwater fishes are significantly threatened by alien species invasions, the presence of dams, and water withdrawals associated with agricultural and urban use. A long history of habitat degradation and dependence of salmonid taxa on hatchery supplementation are also contributing to the decline of fish- es in the state. These threats are exacerbated by climate change, which is also reducing suitable habitats through increases in temperatures and chang- es to flow regimes. Approximately 80% of freshwater fishes are now facing extinction in the next 100 years, unless current trends are reversed by active conservation. Here, we review threats to California freshwater fishes and update a five-tiered approach to preserve aquatic biodiversity in California, with emphasis on fish species diversity. Central to the approach are man- agement actions that address conservation at different scales, from single taxon and species assemblages to Aquatic Diversity Management Areas, wa- tersheds, and bioregions. Keywords: alien fishes, climate change, conservation strategy, dams Citation: Quiñones RM, Moyle PB (2015) California’s freshwater fishes: status and man- agement. -

CATALINA CALIFORNIA QUAIL (Callipepla Californica Catalinensis) Paul W
II SPECIES ACCOUNTS Andy Birch PDF of Catalina California Quail account from: Shuford, W. D., and Gardali, T., editors. 2008. California Bird Species of Special Concern: A ranked assessment of species, subspecies, and distinct populations of birds of immediate conservation concern in California. Studies of Western Birds 1. Western Field Ornithologists, Camarillo, California, and California Department of Fish and Game, Sacramento. California Bird Species of Special Concern CATALINA CALIFORNIA QUAIL (Callipepla californica catalinensis) Paul W. Collins Criteria Scores Population Trend 0 Santa Range Trend 0 Barbara County Population Size 7.5 Range Size 10 Ventura Endemism 10 County Population Concentration 10 Threats 0 Los San Miguel Is. Santa Cruz Is. Angeles County Anacapa Is. Santa Rosa Is. Santa Barbara Is. Santa Catalina Is. San Nicolas Is. San Clemente Is. Current Year-round Range Historic Year-round Range County Boundaries Kilometers 20 10 0 20 Current and historic (ca. 1944) year-round range of the Catalina California Quail. Birds from Santa Catalina Island (perhaps brought by Native Americans) later introduced successfully to Santa Rosa (1935–1940) and Santa Cruz (late 1940s) islands, but unsuccessfully to San Nicolas Island (1962); quail from mainland populations of C. c. californica introduced unsuccessfully to Santa Cruz (prior to 1875) and San Clemente (late 19th century, 1913) islands. Catalina California Quail Studies of Western Birds 1:107–111, 2008 107 Studies of Western Birds No. 1 SPECIAL CONCERN PRIORITY HISTORIC RANGE AND ABUNDANCE Currently considered a Bird Species of Special IN CALIFORNIA Concern (year round), priority 3. This subspecies Grinnell and Miller (1944) described the Catalina was not included on prior special concern lists California Quail as a “common to abundant” (Remsen 1978, CDFG 1992). -

Natural History Museum Unveils Portrait of Juana María, the Lone Woman of San Nicolas Island
Media Contact: Briana Sapp Tivey Director of Marketing and Communications Email: [email protected] Phone: 805-682-4711 ext. 117 FOR IMMEDIATE RELEASE Natural History Museum Unveils Portrait of Juana María, the Lone Woman of San Nicolas Island Local Artist Holli Harmon Creates Likeness Based on First-Hand Accounts Santa Barbara, California (February 21, 2018) — The Santa Barbara Museum of Natural History recently unveiled a historically accurate portrait of Juana María, the Lone Woman of San Nicolas Island. Fictionalized as “Karana” in Scott O’Dell’s novel Island of the Blue Dolphins, she was a real person who lived by herself on San Nicolas Island. Accidentally left behind in 1835, when the last of the native inhabitants were conveyed to the mainland at the request of the Santa Barbara Mission priests, she resourcefully caught her own food, made her own clothes, and built her own shelter for 18 years. In 1853, Carl Dittman and sea otter hunter Captain George Nidever found the woman alive and well. She willingly returned to the mainland on his ship, living with Nidever’s family in Santa Barbara for only seven weeks before she tragically fell ill and died. The Lone Woman was conditionally baptized with the name Juana María and buried at the Santa Barbara Mission. Harmon’s piece is the first painting to be based on historical records. Most representations of Juana María to date have been based on the romantic image popularized in O’Dell’s book. A research team including archaeologist Steve Schwartz, historian Susan Morris, and Museum Curator of Anthropology John Johnson supplied local artist Holli Harmon with historically accurate descriptions of the Lone Woman. -

Rare Artifact Found on San Clemente Island Mass Communication Specialist 2Nd Class Shawnte Bryan July 16, 2013 (U.S
Santa Cruz Archaeological Society Newsletter Rare Artifact Found on San Clemente Island Mass Communication Specialist 2nd Class Shawnte Bryan July 16, 2013 (U.S. Navy Story Number: NNS130716-29) The Navy discovered a significant prehistoric artifact 90 miles west of San Diego on San Clemente Island (SCI) located mid-island at a newly discovered archeological site. A boat effigy made of submarine volcano lava was spotted at the surface of the site during an archeology survey. The boat effigy represents a type of boat used by the California Indians who occupied the California Channels and adjacent southern California mainland at the time of the Spanish "discovery" in the AD 1500s. Dr. Andy Yatsko, Senior Archaeologist and Region Southwest Archaeologist for the Naval Facilities Engineering Command Southwest in San Diego, who has over 40 years experience in prehistoric and historic archaeology finds the boat effigy to be an uncommon find. "Boat effigies like the one found are exceedingly rare in the archaeological record, with this being my first one recovered during my 30 year tenure with SCI," said Dr. Yatsko [image courtesy U.S. Navy]. "Finding artifacts on the surface of archaeological sites at the island is not unusual, but a rare one like this is always exciting to come across." The effigy was created from submarine volcano lava. This lava differs from the lava that flows on the mainland because the vesicles are smaller, making it more brittle and more difficult to handle. “The vesicle volcanic material used is hard and brittle but somehow they were able to craft a fine little carving out of it to represent an important part of their culture," said Yatsko. -

UNIVERSITY of CALIFORNIA Santa Barbara Ancient Plant Use and the Importance of Geophytes Among the Island Chumash of Santa Cruz
UNIVERSITY OF CALIFORNIA Santa Barbara Ancient Plant Use and the Importance of Geophytes among the Island Chumash of Santa Cruz Island, California A dissertation submitted in partial satisfaction of the requirements for the degree of Doctor of Philosophy in Anthropology by Kristina Marie Gill Committee in charge: Professor Michael A. Glassow, Chair Professor Michael A. Jochim Professor Amber M. VanDerwarker Professor Lynn H. Gamble September 2015 The dissertation of Kristina Marie Gill is approved. __________________________________________ Michael A. Jochim __________________________________________ Amber M. VanDerwarker __________________________________________ Lynn H. Gamble __________________________________________ Michael A. Glassow, Committee Chair July 2015 Ancient Plant Use and the Importance of Geophytes among the Island Chumash of Santa Cruz Island, California Copyright © 2015 By Kristina Marie Gill iii DEDICATION This dissertation is dedicated to my Family, Mike Glassow, and the Chumash People. iv ACKNOWLEDGEMENTS I am indebted to many people who have provided guidance, encouragement, and support in my career as an archaeologist, and especially through my undergraduate and graduate studies. For those of whom I am unable to personally thank here, know that I deeply appreciate your support. First and foremost, I want to thank my chair Michael Glassow for his patience, enthusiasm, and encouragement during all aspects of this daunting project. I am also truly grateful to have had the opportunity to know, learn from, and work with my other committee members, Mike Jochim, Amber VanDerwarker, and Lynn Gamble. I cherish my various field experiences with them all on the Channel Islands and especially in southern Germany with Mike Jochim, whose worldly perspective I value deeply. I also thank Terry Jones, who provided me many undergraduate opportunities in California archaeology and encouraged me to attend a field school on San Clemente Island with Mark Raab and Andy Yatsko, an experience that left me captivated with the islands and their history. -
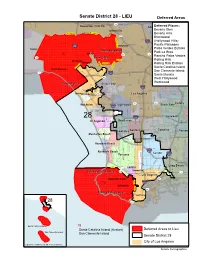
Assigned Deferred Area Map 2
Senate District 28 - LIEU Deferred Areas d C n a 7 Ve a n r 2 t n ura e v B o Moorpark 134 li g ST O 159 Colorado a ST Sherman Oaks Studio City DeferCred Places: ol Gllendalle V ora 134 e do ST r 710 d ST d Bevu erly Glen 0 Universal City n g 1 a o r Mulh Mulh d F 1 olland oll B nd Dr and an i Mulholla Dr l g l Beverly Hills u o e lh r Mu 101 York o a N ¤£ i n c z Brentwood h o l i o y Fe l s n o s L 2 a ST C R C i a v (Hollywood Hills) 0 e r n 5 y 7 rs e y e 1 id t r B o § ¨ e a ¦ e ood n Hollyw t e C n v w R S e Paciyfic Palisaodes e d t k p l unse d Hollywood u 405 S n r M r o a e 4 ly s R s 3 C e L s ¦¨§ G t r iv e e le v Palor s Verdes Estates t Topanga il s S n m i d n S u e e i o d p West Hollywood 110 West Hollywood m t Melrose a u t ST e e l s SPark La2 Brea r v 6 n F e Bel Air u d S Beverly 2 16 a Sun x 170 ST s a ST e f t r i g Rancrinho Palos Verdes a 3rd p 27 F ST Beverrlly Hiilllls S Va W 6th St lley Wilshire RT olling Hills Brentwood H e e e v m 10 a A 6 p th Westwood t Whitworth Dr le x ¨§ h ¦ e t v a A f Rolling Hills Estates A r e v i s a e 10 un n a 1 S a F st t n 8 ¨§ Pico ¦ n t o o S h s a M t Santa Catalina Island ic r e avez p r V Cesar E Ch t e e 7 4 m B n t t e Pacific Palisades y b i h h s 2 l ce 6 o a n O th B r L n u R u to o n t San Clemente Island S o dy g n 1 M n i r ST 10 l r a 60 e n l t a A ST ta 2 n § s ¨ Sante a Monica o ¦ a n ti 5 7 o a F 2 E M 20 N a ¨§ Rodeo ¦ th 187 t ST n t S a n West Hollywood o S o t l o n i m 7 a r r 8 a 1 t 710 e M n V B e Westwood an ¨§ d ¦ er C in Santta Moniica 87 Culver -

ASSESSMENT of COASTAL WATER RESOURCES and WATERSHED CONDITIONS at CHANNEL ISLANDS NATIONAL PARK, CALIFORNIA Dr. Diana L. Engle
National Park Service U.S. Department of the Interior Technical Report NPS/NRWRD/NRTR-2006/354 Water Resources Division Natural Resource Program Centerent of the Interior ASSESSMENT OF COASTAL WATER RESOURCES AND WATERSHED CONDITIONS AT CHANNEL ISLANDS NATIONAL PARK, CALIFORNIA Dr. Diana L. Engle The National Park Service Water Resources Division is responsible for providing water resources management policy and guidelines, planning, technical assistance, training, and operational support to units of the National Park System. Program areas include water rights, water resources planning, marine resource management, regulatory guidance and review, hydrology, water quality, watershed management, watershed studies, and aquatic ecology. Technical Reports The National Park Service disseminates the results of biological, physical, and social research through the Natural Resources Technical Report Series. Natural resources inventories and monitoring activities, scientific literature reviews, bibliographies, and proceedings of technical workshops and conferences are also disseminated through this series. Mention of trade names or commercial products does not constitute endorsement or recommendation for use by the National Park Service. Copies of this report are available from the following: National Park Service (970) 225-3500 Water Resources Division 1201 Oak Ridge Drive, Suite 250 Fort Collins, CO 80525 National Park Service (303) 969-2130 Technical Information Center Denver Service Center P.O. Box 25287 Denver, CO 80225-0287 Cover photos: Top Left: Santa Cruz, Kristen Keteles Top Right: Brown Pelican, NPS photo Bottom Left: Red Abalone, NPS photo Bottom Left: Santa Rosa, Kristen Keteles Bottom Middle: Anacapa, Kristen Keteles Assessment of Coastal Water Resources and Watershed Conditions at Channel Islands National Park, California Dr. Diana L. -

Chumash Ritual and Sacred Geography on Santa Cruz Island, California
UC Merced Journal of California and Great Basin Anthropology Title Chumash Ritual and Sacred Geography on Santa Cruz Island, California Permalink https://escholarship.org/uc/item/0z15r2hj Journal Journal of California and Great Basin Anthropology, 27(2) ISSN 0191-3557 Author Perry, Jennifer E Publication Date 2007 Peer reviewed eScholarship.org Powered by the California Digital Library University of California Journal of California and Great Basin Anthropology | Vol, 27, No, 2 (2007) | pp. 103-124 Chumash Ritual and Sacred Geography on Santa Cruz Island, California JENNIFER E. PERRY Department of Anthropology, Pomona College, Claremont, CA 91711 In contrast to the archaeological visibility of Chumash rock art on the mainland, its virtual absence on the northern Channel Islands is reflective of what little is understood about ritual behavior in island prehistory. By relying on relevant ethnohistoric and ethnographic references from the mainland, it is possible to evaluate how related activities may be manifested archaeologically on the islands. On Santa Cruz Island, portable ritual items and rock features have been identified on El Montahon and the North Ridge, the most prominent ridgelines on the northern islands Citing material correlates of ritual behavior, intentionally-made rock features are interpreted as possible shrines, which were an important aspect of winter solstice ceremonies among the mainland Chumash. Portable ritual items and possible shrines are considered in the context of sacred geography, revealing aspects of how the Chumash may have interacted with the supernatural landscape of Santa Cruz Island. andscapes are imbued with different attributes and Conception and Mount Pinos (as examples of the former) Llvalues; whether economic, aesthetic, recrea to sweatlodges and rock sites (as examples of the latter) tional, spiritual, or otherwise, these values intersect, (Grant 1965; Haley and Wilcoxon 1997,1999). -

Species at Risk on Department of Defense Installations
Species at Risk on Department of Defense Installations Revised Report and Documentation Prepared for: Department of Defense U.S. Fish and Wildlife Service Submitted by: January 2004 Species at Risk on Department of Defense Installations: Revised Report and Documentation CONTENTS 1.0 Executive Summary..........................................................................................iii 2.0 Introduction – Project Description................................................................. 1 3.0 Methods ................................................................................................................ 3 3.1 NatureServe Data................................................................................................ 3 3.2 DOD Installations............................................................................................... 5 3.3 Species at Risk .................................................................................................... 6 4.0 Results................................................................................................................... 8 4.1 Nationwide Assessment of Species at Risk on DOD Installations..................... 8 4.2 Assessment of Species at Risk by Military Service.......................................... 13 4.3 Assessment of Species at Risk on Installations ................................................ 15 5.0 Conclusion and Management Recommendations.................................... 22 6.0 Future Directions............................................................................................. -

How to Look at Oaks
How to Look at Oaks Species name: __________________________________________ Growth habit: tree or shrub Bark type on mature trees: scaly and papery or smooth and furrowed gray or black Foliage: Evergreen or deciduous Leaves Leaf margins: entire (smooth) lobed toothed Leaf edges from the side: wavy fat concave Lobe tips: rounded (not spiny) or spine-tipped Leaf underside: hairless or with small tufs of hairs or covered with hairs color: _______________ Leaf upper surface: dull or shiny hairless or covered with hairs color: _______________ Acorns Time to maturity: one year or two years Acorn shape: oblong elongated round tip: pointed or rounded Acorn size: length: ___________ width: ___________ Acorn cup: deep or shallow % of mature acorn covered by cup: Acorn cup shape: cap saucer bowl cup Acorn cup scales: thin, papery, leafike or thick, knobby, warty Scale tips: loose or pressed tightly to each other Acorn inner surface: densely fuzzy or hairless Based on the features above, is this a: Red/Black Oak White Oak Intermediate Oak Other Notable Features: Characteristics and Taxonomy of Quercus in California Genus Quercus = ~400-600 species Original publication: Linnaeus, Species Plantarum 2: 994. 1753 Sections in the subgenus Quercus: Red Oaks or Black Oaks 1. Foliage evergreen or deciduous (Quercus section Lobatae syn. 2. Mature bark gray to dark brown or black, smooth or Erythrobalanus) deeply furrowed, not scaly or papery ~195 species 3. Leaf blade lobes with bristles 4. Acorn requiring 2 seasons to mature (except Q. Example native species: agrifolia) kelloggii, agrifolia, wislizeni, parvula 5. Cup scales fattened, never knobby or warty, never var. -

Classification of the House Finch of the Channel Islands, Southern California Philip Unitt, San Diego Natural History Museum, P
NOTES CLASSIFICATION OF THE HOUSE FINCH OF THE CHANNEL ISLANDS, SOUTHERN CALIFORNIA PHILIP UNITT, San Diego Natural History Museum, P. O. Box 121390, San Diego, California 92112-1390; [email protected] The Channel Islands of southern California are renowned for their many endemic plants and animals. Among land birds, 22 endemic species or subspecies have been described. Not all of these are valid, however, and their distinctiveness spans a wide spectrum. Johnson (1972) categorized the islands’ land birds in five strata ranging from the Island Scrub-Jay (Aphelocoma insularis) of Santa Cruz Island to those not known to differ from mainland populations. The last includes both species of which no subspecies endemic to the islands have been proposed, such as the Bushtit (Psaltriparus minimus), and described subspecies whose supposed differences have been discredited or found to be insufficient to meet the criteria for recognition. These synonymized names include Colaptes cafer sedentarius van Rossem, 1944 (with C. auratus collaris; AOU 1952), Vireo mailliardorum Grinnell, 1903 (with V. h. huttoni; AOU 1908), Salpinctes obsoletus pulverius Grinnell, 1898 (with S. o. obsoletus; Grinnell 1929), Amphispiza belli clementeae Ridgway, 1898 (with A. b. belli; Patten and Unitt 2002), and Melospiza melodia micronyx and M. m. clementae (the last two with M. m. graminea; Patten and Pruett 2009). The status of the supposed subspecies of the House Finch (Carpodacus mexicanus clementis) has also been questioned. In the original description, Mearns (1898:259) reported the House Finches of the Channel Islands to differ from those of the mainland (C. m. frontalis) in their larger legs and feet, broader streaking on the underparts, shorter wings, and “bill much larger and more convex above.” He designated a type specimen from San Clemente Island, U.S. -

References.Qxd 12/14/2004 10:35 AM Page 771
Ducks_References.qxd 12/14/2004 10:35 AM Page 771 References Aarvak, T. and Øien, I.J. 1994. Dverggås Anser Adams, J.S. 1971. Black Swan at Lake Ellesmere. erythropus—en truet art i Norge. Vår Fuglefauna 17: 70–80. Wildl. Rev. 3: 23–25. Aarvak, T. and Øien, I.J. 2003. Moult and autumn Adams, P.A., Robertson, G.J. and Jones, I.L. 2000. migration of non-breeding Fennoscandian Lesser White- Time-activity budgets of Harlequin Ducks molting in fronted Geese Anser erythropus mapped by satellite the Gannet Islands, Labrador. Condor 102: 703–08. telemetry. Bird Conservation International 13: 213–226. Adrian, W.L., Spraker, T.R. and Davies, R.B. 1978. Aarvak, T., Øien, I.J. and Nagy, S. 1996. The Lesser Epornitics of aspergillosis in Mallards Anas platyrhynchos White-fronted Goose monitoring programme,Ann. Rept. in north central Colorado. J. Wildl. Dis. 14: 212–17. 1996, NOF Rappportserie, No. 7. Norwegian Ornitho- AEWA 2000. Report on the conservation status of logical Society, Klaebu. migratory waterbirds in the agreement area. Technical Series Aarvak, T., Øien, I.J., Syroechkovski Jr., E.E. and No. 1.Wetlands International,Wageningen, Netherlands. Kostadinova, I. 1997. The Lesser White-fronted Goose Afton, A.D. 1983. Male and female strategies for Monitoring Programme.Annual Report 1997. Klæbu, reproduction in Lesser Scaup. Unpubl. Ph.D. thesis. Norwegian Ornithological Society. NOF Raportserie, Univ. North Dakota, Grand Forks, US. Report no. 5-1997. Afton, A.D. 1984. Influence of age and time on Abbott, C.C. 1861. Notes on the birds of the Falkland reproductive performance of female Lesser Scaup.