Feeding by Euplotes Mutabilis (Ciliophora, Hypotrichida)*
Total Page:16
File Type:pdf, Size:1020Kb

Load more
Recommended publications
-
Molecular Data and the Evolutionary History of Dinoflagellates by Juan Fernando Saldarriaga Echavarria Diplom, Ruprecht-Karls-Un
Molecular data and the evolutionary history of dinoflagellates by Juan Fernando Saldarriaga Echavarria Diplom, Ruprecht-Karls-Universitat Heidelberg, 1993 A THESIS SUBMITTED IN PARTIAL FULFILMENT OF THE REQUIREMENTS FOR THE DEGREE OF DOCTOR OF PHILOSOPHY in THE FACULTY OF GRADUATE STUDIES Department of Botany We accept this thesis as conforming to the required standard THE UNIVERSITY OF BRITISH COLUMBIA November 2003 © Juan Fernando Saldarriaga Echavarria, 2003 ABSTRACT New sequences of ribosomal and protein genes were combined with available morphological and paleontological data to produce a phylogenetic framework for dinoflagellates. The evolutionary history of some of the major morphological features of the group was then investigated in the light of that framework. Phylogenetic trees of dinoflagellates based on the small subunit ribosomal RNA gene (SSU) are generally poorly resolved but include many well- supported clades, and while combined analyses of SSU and LSU (large subunit ribosomal RNA) improve the support for several nodes, they are still generally unsatisfactory. Protein-gene based trees lack the degree of species representation necessary for meaningful in-group phylogenetic analyses, but do provide important insights to the phylogenetic position of dinoflagellates as a whole and on the identity of their close relatives. Molecular data agree with paleontology in suggesting an early evolutionary radiation of the group, but whereas paleontological data include only taxa with fossilizable cysts, the new data examined here establish that this radiation event included all dinokaryotic lineages, including athecate forms. Plastids were lost and replaced many times in dinoflagellates, a situation entirely unique for this group. Histones could well have been lost earlier in the lineage than previously assumed. -

Suitability of Great South Bay, New York to Blooms of Pfiesteria Piscicida and P
City University of New York (CUNY) CUNY Academic Works School of Arts & Sciences Theses Hunter College Summer 8-10-2015 Suitability of Great South Bay, New York to Blooms of Pfiesteria piscicida and P. shumwayae Prior to Superstorm Sandy, October 29, 2012 Pawel Tomasz Zablocki CUNY Hunter College How does access to this work benefit ou?y Let us know! More information about this work at: https://academicworks.cuny.edu/hc_sas_etds/6 Discover additional works at: https://academicworks.cuny.edu This work is made publicly available by the City University of New York (CUNY). Contact: [email protected] Suitability of Great South Bay, New York, to Blooms of Pfiesteria piscicida and P. shumwayae Prior to Superstorm Sandy, October 29, 2012. By Pawel Zablocki Submitted in partial fulfillment of the requirements for the degree of Master of Arts Hunter College of the City of New York 2015 Thesis sponsor: __25 July 2015 Peter X. Marcotullio Date First Reader _2 August 2015 Karl H. Szekielda Date Second Reader i Acknowledgements I would like to thank my advisor, Professor H. Gong and two of my excellent readers—Professor Peter Marcotullio and Professor Karl Szekielda who provided their invaluable advice, alleviated my concerns, and weathered the avalanche of my questions. ii Abstract of the Thesis Pfiesteria piscicida and P. shumwayae are toxic dinoflagellates implicated in massive fish kills in North Carolina and Maryland during 1990s. A set of physical, chemical, and biological factors influence population dynamics of these organisms. This study employs information gathered from relevant literature on temperature, salinity, dissolved oxygen, pH, turbulent mixing, and dissolved nutrients, bacteria, algae, microzooplankton, mesozooplankton, bivalve mollusks, finfish, and other toxic dinoflagellates, which influence Pfiesteria population dynamics. -

University of Oklahoma
UNIVERSITY OF OKLAHOMA GRADUATE COLLEGE MACRONUTRIENTS SHAPE MICROBIAL COMMUNITIES, GENE EXPRESSION AND PROTEIN EVOLUTION A DISSERTATION SUBMITTED TO THE GRADUATE FACULTY in partial fulfillment of the requirements for the Degree of DOCTOR OF PHILOSOPHY By JOSHUA THOMAS COOPER Norman, Oklahoma 2017 MACRONUTRIENTS SHAPE MICROBIAL COMMUNITIES, GENE EXPRESSION AND PROTEIN EVOLUTION A DISSERTATION APPROVED FOR THE DEPARTMENT OF MICROBIOLOGY AND PLANT BIOLOGY BY ______________________________ Dr. Boris Wawrik, Chair ______________________________ Dr. J. Phil Gibson ______________________________ Dr. Anne K. Dunn ______________________________ Dr. John Paul Masly ______________________________ Dr. K. David Hambright ii © Copyright by JOSHUA THOMAS COOPER 2017 All Rights Reserved. iii Acknowledgments I would like to thank my two advisors Dr. Boris Wawrik and Dr. J. Phil Gibson for helping me become a better scientist and better educator. I would also like to thank my committee members Dr. Anne K. Dunn, Dr. K. David Hambright, and Dr. J.P. Masly for providing valuable inputs that lead me to carefully consider my research questions. I would also like to thank Dr. J.P. Masly for the opportunity to coauthor a book chapter on the speciation of diatoms. It is still such a privilege that you believed in me and my crazy diatom ideas to form a concise chapter in addition to learn your style of writing has been a benefit to my professional development. I’m also thankful for my first undergraduate research mentor, Dr. Miriam Steinitz-Kannan, now retired from Northern Kentucky University, who was the first to show the amazing wonders of pond scum. Who knew that studying diatoms and algae as an undergraduate would lead me all the way to a Ph.D. -

The Intestinal Protozoa
The Intestinal Protozoa A. Introduction 1. The Phylum Protozoa is classified into four major subdivisions according to the methods of locomotion and reproduction. a. The amoebae (Superclass Sarcodina, Class Rhizopodea move by means of pseudopodia and reproduce exclusively by asexual binary division. b. The flagellates (Superclass Mastigophora, Class Zoomasitgophorea) typically move by long, whiplike flagella and reproduce by binary fission. c. The ciliates (Subphylum Ciliophora, Class Ciliata) are propelled by rows of cilia that beat with a synchronized wavelike motion. d. The sporozoans (Subphylum Sporozoa) lack specialized organelles of motility but have a unique type of life cycle, alternating between sexual and asexual reproductive cycles (alternation of generations). e. Number of species - there are about 45,000 protozoan species; around 8000 are parasitic, and around 25 species are important to humans. 2. Diagnosis - must learn to differentiate between the harmless and the medically important. This is most often based upon the morphology of respective organisms. 3. Transmission - mostly person-to-person, via fecal-oral route; fecally contaminated food or water important (organisms remain viable for around 30 days in cool moist environment with few bacteria; other means of transmission include sexual, insects, animals (zoonoses). B. Structures 1. trophozoite - the motile vegetative stage; multiplies via binary fission; colonizes host. 2. cyst - the inactive, non-motile, infective stage; survives the environment due to the presence of a cyst wall. 3. nuclear structure - important in the identification of organisms and species differentiation. 4. diagnostic features a. size - helpful in identifying organisms; must have calibrated objectives on the microscope in order to measure accurately. -

Parameterisation of Microprotozooplankton Grazing and Growth
Parameterisation of Microprotozooplankton Grazing and Growth: From data analysis to simulations in ecosystem model coupled to general circulation-biogeochemical model. Dissertation Zur Erlangung des Akademischen Grades eines Doktors der Naturwissenschaften - Dr. rer. Nat. – Im Fachbereich 2 (Biologie/Chemie) Der Universität Bremen vorgelegt von Sévrine Sailley Institutes: Alfred Wegener Institut for Polar and Marine Research (AWI), Bremerhaven, Germany University of East Anglia (UEA), Norwich, UK British Antarctic Survey (BAS), Cambridge, UK University Bremen, Germany This work was funded by Euroceans program, project number WP3.2-SYS-1092. 2 Erster Gutachter: Prof. Dr. Dieter Wolf-Gladrow Zweiter Gutachter: Prof. Dr. Corinne Le Quéré Tag des öffentlichen Kolloquims: Universität Bremen, 23 November 2009 Eidesstattliche Erklärung Hiermit erkläre ich nach § 6 Abs. 5 der Promotionsordnung der Uni Bremen (vom 14 März 2007), dass ich die Vorliegende Dissertation (1) ohne unerlaubte Hilfe angefertigt habe, (2) keine anderen als die von mir angegebenen Quellen und Hilfsmittel benutzt habe und (3) die den benutzen Werken wörtlich oder inhaltlich entnommen Stellen als solche kenntlich gemacht habe Sévrine Sailley 3 4 Acknowledgments Here, I would like to thank people who helped me in different ways through this thesis. There are quite a lot of people who should figure here, so if I’ve forgotten somebody, I’m sorry it wasn’t on purpose. Thanks to Christine and Dieter for welcoming me at AWI and in Bremerhaven and helping not just with the thesis but also simply life in Germany, thanks for being more than just supervisors. Thanks to Clare Enright for helping me arrange my stay at BAS, but also with going through all the bugs that happened in PlankTOM, or simply little problems with the cluster. -

Phylogenomic Analysis of Balantidium Ctenopharyngodoni (Ciliophora, Litostomatea) Based on Single-Cell Transcriptome Sequencing
Parasite 24, 43 (2017) © Z. Sun et al., published by EDP Sciences, 2017 https://doi.org/10.1051/parasite/2017043 Available online at: www.parasite-journal.org RESEARCH ARTICLE Phylogenomic analysis of Balantidium ctenopharyngodoni (Ciliophora, Litostomatea) based on single-cell transcriptome sequencing Zongyi Sun1, Chuanqi Jiang2, Jinmei Feng3, Wentao Yang2, Ming Li1,2,*, and Wei Miao2,* 1 Hubei Key Laboratory of Animal Nutrition and Feed Science, Wuhan Polytechnic University, Wuhan 430023, PR China 2 Institute of Hydrobiology, Chinese Academy of Sciences, No. 7 Donghu South Road, Wuchang District, Wuhan 430072, Hubei Province, PR China 3 Department of Pathogenic Biology, School of Medicine, Jianghan University, Wuhan 430056, PR China Received 22 April 2017, Accepted 12 October 2017, Published online 14 November 2017 Abstract- - In this paper, we present transcriptome data for Balantidium ctenopharyngodoni Chen, 1955 collected from the hindgut of grass carp (Ctenopharyngodon idella). We evaluated sequence quality and de novo assembled a preliminary transcriptome, including 43.3 megabits and 119,141 transcripts. Then we obtained a final transcriptome, including 17.7 megabits and 35,560 transcripts, by removing contaminative and redundant sequences. Phylogenomic analysis based on a supermatrix with 132 genes comprising 53,873 amino acid residues and phylogenetic analysis based on SSU rDNA of 27 species were carried out herein to reveal the evolutionary relationships among six ciliate groups: Colpodea, Oligohymenophorea, Litostomatea, Spirotrichea, Hetero- trichea and Protocruziida. The topologies of both phylogenomic and phylogenetic trees are discussed in this paper. In addition, our results suggest that single-cell sequencing is a sound method of obtaining sufficient omics data for phylogenomic analysis, which is a good choice for uncultivable ciliates. -

The Life Cycle of Trypanosoma Cruzi
Tyler et al. THE LIFE CYCLE OF TRYPANOSOMA CRUZI K. M. Tyler, C. L. Olson and D. M. Engman Departments of Microbiology-Immunology and Pathology Feinberg Medical School of Northwestern University, Chicago, IL 60611 ABSTRACT Since the discovery of Trypanosoma cruzi as the parasite that causes Chagas disease, nearly a century ago, the details of the organism's life cycle have fascinated scientists. T. cruzi is a single-celled eukaryote with a complex life cycle alternating between reduviid bug vectors and vertebrate hosts. It is able to adapt via the process of cellular differentiation to replicate within the diverse environments represented of the insect's gut and host cell cytoplasm. These adaptive transformations take the form of coordinated changes in morphology, metabolism and cell cycle regulation. Different life cycle stages of T. cruzi show dramatically different protein and RNA profiles, which are the end result of unusual mechanisms for regulating gene expression. In recent years, new molecular techniques have been brought to bear on the life cycle dramatically increasing our knowledge of the strategies employed by the parasite to ensure its continued survival. INTRODUCTION Chagas disease The etiologic agent of the chronic and often fatal Chagas disease is the American trypanosome, Trypanosoma cruzi , a flagellated protozoan of the order Kinetoplastida. The survival of T. cruzi is dependent on the successful transmission between, and the colonization of, two radically different environments: the midgut of the reduviid bug vector and the cytoplasm of the mammalian host cell. As is true of all infections, interruption of the pathogen's life cycle will lead to eradication of the disease. -

A PRELIMINARY NOTE on TETRAMITUS, a STAGE in the LIFE CYCLE of a COPROZOIC AMOEBA by MARTHA BUNTING
294 Z6OL6G Y.- M. BUNTING PROC. N. A. S. A PRELIMINARY NOTE ON TETRAMITUS, A STAGE IN THE LIFE CYCLE OF A COPROZOIC AMOEBA By MARTHA BUNTING DEPARTMENT OF ZOOLOGY, UNIVERSITY OF PENNSYLVANIA Communicated July 24, 1922 In 1920 a study of the Entamoeba of the rat was begun and in a culture on artificial medium a number of Coprozoic amoeba appeared, one of which exhibited a flagellate phase which seems to be identical with Tetramitus rostratus Perty.1 The appearance of flagellate phases in cultures of Coprozoic amoeba (Whitmore,2 etc.) as well as of soil amoeba (Kofoid,1 Wilson,4 etc.) has been recorded a number of times, but the flagellates which appeared were relatively simple in organization and very transitory in occurrence. Fur- thermore, some of the simpler flagellates have been observed (Pascher5) to lose their flagella and become amoeboid. In the case here described, however, there is a transformation of a simple amoeba into a relatively complex flagellate which multiplies for several days then returns to the amoeboid condition and becomes encysted. Tetramitus rostratus has been studied by a number of investigators since its discovery in 1852, by Perty,' yet no one has given a full account of its life history. The lack of cysts in previous accounts, mentioned by Dobell and O'Connor,6 is explained by the transformation into an amoeba before encystment. It is believed that this animal presents the extreme in transformations of this sort and serves to emphasize the close relationship between amoebae and flagellates, and the need for careful studies of life cycles, in pure cultures where possible, in investigations of coprozoic and other Protozoa. -

Grazing of Two Euplotid Ciliates on the Heterotrophic Dinoflagellates Pfiesteria Piscicida and Cryptoperidiniopsis Sp
AQUATIC MICROBIAL ECOLOGY Vol. 33: 303–308, 2003 Published November 7 Aquat Microb Ecol NOTE Grazing of two euplotid ciliates on the heterotrophic dinoflagellates Pfiesteria piscicida and Cryptoperidiniopsis sp. Scott G. Gransden1, Alan J. Lewitus1, 2,* 1Belle W. Baruch Institute for Marine and Coastal Science, University of South Carolina, PO Box 1630, Georgetown, South Carolina 29442, USA 2Marine Resources Research Institute, SC Department of Natural Resources, Hollings Marine Laboratory, 331 Fort Johnson Road, Charleston, South Carolina 29412, USA ABSTRACT: Pfiesteria piscicida and Cryptoperidiniopsis spp. breaks have been associated with millions of dollars of are common co-occurring heterotrophic dinoflagellates in lost revenue to the fisheries and tourism industries estuaries along the Atlantic coast of the United States. We (Burkholder & Glasgow 1997, CENR 2000). isolated P. piscicida, Cryptoperidiniopsis sp., and 2 benthic ciliates (Euplotes vannus and E. woodruffi) from North Inlet Increased awareness of Pfiesteria spp.’s potential estuary, South Carolina, and examined the growth and graz- impact on environmental and human health has led ing properties of the ciliates on cultures of the dinoflagellates to several studies on the dinoflagellates’ trophic dy- maintained with cryptophyte (Storeatula major) prey. Ciliate namics. Ingestion of P. piscicida by copepods, rotifers, growth and grazing parameters on cryptophyte monocultures or benthic ciliates has been reported (Burkholder & and mixed diets of cryptophytes and P. piscicida were signifi- cantly higher with E. woodruffi than E. vannus. Also, the net Glasgow 1995, Mallin et al. 1995). More recently, grazing impact of E. woodruffi on P. piscicida prey was higher Stoecker et al. (2000) added 5-chloromethylfluorescein than the impact on Cryptoperidiniopsis sp., while the E. -

A Monograph on the Protozoa of the Large Intestine of the Horse Ta-Shih Hsiung Iowa State College
Iowa State University Capstones, Theses and Retrospective Theses and Dissertations Dissertations 1930 A monograph on the protozoa of the large intestine of the horse Ta-Shih Hsiung Iowa State College Follow this and additional works at: https://lib.dr.iastate.edu/rtd Part of the Zoology Commons Recommended Citation Hsiung, Ta-Shih, "A monograph on the protozoa of the large intestine of the horse" (1930). Retrospective Theses and Dissertations. 14768. https://lib.dr.iastate.edu/rtd/14768 This Dissertation is brought to you for free and open access by the Iowa State University Capstones, Theses and Dissertations at Iowa State University Digital Repository. It has been accepted for inclusion in Retrospective Theses and Dissertations by an authorized administrator of Iowa State University Digital Repository. For more information, please contact [email protected]. INFORMATION TO USERS This manuscript has been reproduced from the microfilm master. UMI films the text directly from the original or copy submitted. Thus, some thesis and dissertation copies are in typewriter face, while others may be from any type of computer printer. The quality of this reproduction is dependent upon the quality of the copy submitted. Broken or indistinct print, colored or poor quality illustrations and photographs, print bleedthrough, substandard margins, and improper alignment can adversely affect reproduction. In the unlikely event that the author did not send UMI a complete manuscript and there are missing pages, these will be noted. Also, if unauthorized copyright material had to be removed, a note will indicate the deletion. Oversize materials (e.g., maps, drawings, charts) are reproduced by sectioning the original, beginning at the upper left-hand comer and continuing from left to right in equal sections with small overiaps. -
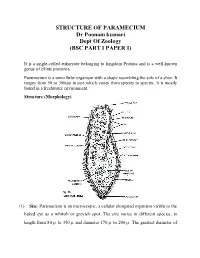
STRUCTURE of PARAMECIUM Dr Poonam Kumari Dept of Zoology (BSC PART I PAPER I)
STRUCTURE OF PARAMECIUM Dr Poonam kumari Dept Of Zoology (BSC PART I PAPER I) It is a single-celled eukaryote belonging to kingdom Protista and is a well-known genus of ciliate protozoa. Paramecium is a unicellular organism with a shape resembling the sole of a shoe. It ranges from 50 to 300um in size which varies from species to species. It is mostly found in a freshwater environment. Structure (Morphology) (1) Size: Paramecium is an microscopic, a cellular elongated organism visible to the baked eye as a whitish or greyish spot. The size varies in different species, in length from 80 to 350 and diameter 170 to 290 . The greatest diameter of the cylindrical body is about two third of its entire length. Usually the individuals of the same species may show minor morphological and physiological differences. (2) Shape: Paramecium is a slipper shaped, cigar shaped, or spindle shaped animalcule. Its shape is usually constant and a symmetrical, because slipper like shape. The body is elongated, blunt and rounded at the anterior end and somewhat pointed of the posterior end. In cross section it is circular with greatest diameter behind the centre of body. The anterior half of the body is slightly twisted. The body is distinguished into an oral or ventral surface and an aboral or dorsal surface. The structure is more complicated due to the development of certain organelles in the acellular body. (3) Oral groove: The ventral surface of body bears a prominent, oblique and shallow depression is called oral groove, it arise from the middle of body and extends to the left side of anterior end. -

Scrippsiella Trochoidea (F.Stein) A.R.Loebl
MOLECULAR DIVERSITY AND PHYLOGENY OF THE CALCAREOUS DINOPHYTES (THORACOSPHAERACEAE, PERIDINIALES) Dissertation zur Erlangung des Doktorgrades der Naturwissenschaften (Dr. rer. nat.) der Fakultät für Biologie der Ludwig-Maximilians-Universität München zur Begutachtung vorgelegt von Sylvia Söhner München, im Februar 2013 Erster Gutachter: PD Dr. Marc Gottschling Zweiter Gutachter: Prof. Dr. Susanne Renner Tag der mündlichen Prüfung: 06. Juni 2013 “IF THERE IS LIFE ON MARS, IT MAY BE DISAPPOINTINGLY ORDINARY COMPARED TO SOME BIZARRE EARTHLINGS.” Geoff McFadden 1999, NATURE 1 !"#$%&'(&)'*!%*!+! +"!,-"!'-.&/%)$"-"!0'* 111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111 2& ")3*'4$%/5%6%*!+1111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111 7! 8,#$0)"!0'*+&9&6"*,+)-08!+ 111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111 :! 5%*%-"$&0*!-'/,)!0'* 11111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111 ;! "#$!%"&'(!)*+&,!-!"#$!'./+,#(0$1$!2! './+,#(0$1$!-!3+*,#+4+).014!1/'!3+4$0&41*!041%%.5.01".+/! 67! './+,#(0$1$!-!/&"*.".+/!1/'!4.5$%"(4$! 68! ./!5+0&%!-!"#$!"#+*10+%,#1$*10$1$! 69! "#+*10+%,#1$*10$1$!-!5+%%.4!1/'!$:"1/"!'.;$*%."(! 6<! 3+4$0&41*!,#(4+)$/(!-!0#144$/)$!1/'!0#1/0$! 6=! 1.3%!+5!"#$!"#$%.%! 62! /0+),++0'* 1111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111<=!