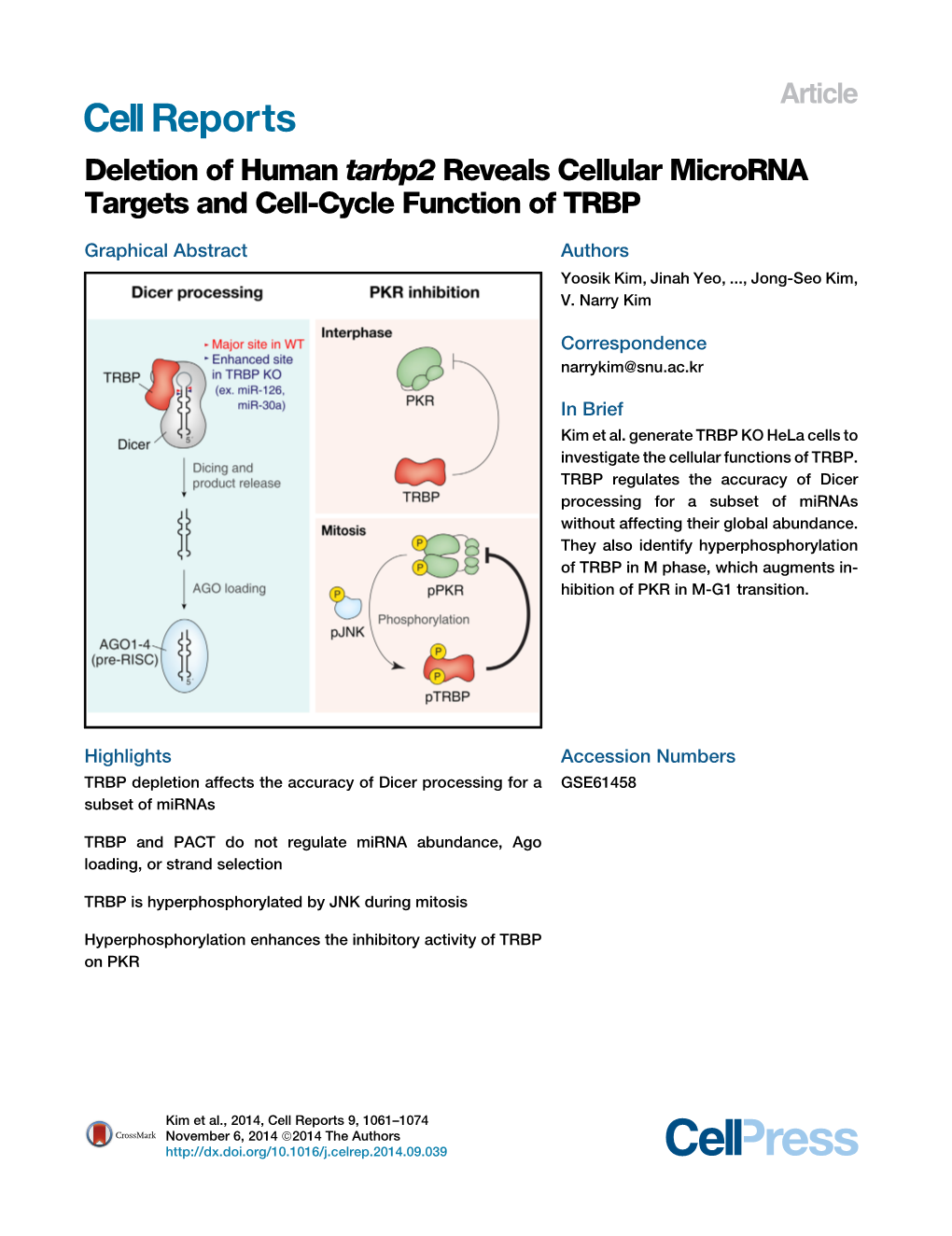
Deletion of Human Tarbp2 Reveals Cellular Microrna Targets and Cell-Cycle Function of TRBP
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Mir-125 in Normal and Malignant Hematopoiesis
Leukemia (2012) 26, 2011–2018 & 2012 Macmillan Publishers Limited All rights reserved 0887-6924/12 www.nature.com/leu SPOTLIGHT REVIEW MiR-125 in normal and malignant hematopoiesis L Shaham1,2, V Binder3,4,NGefen1,5, A Borkhardt3 and S Izraeli1,5 MiR-125 is a highly conserved microRNA throughout many different species from nematode to humans. In humans, there are three homologs (hsa-miR-125b-1, hsa-miR-125b-2 and hsa-miR-125a). Here we review a recent research on the role of miR-125 in normal and malignant hematopoietic cells. Its high expression in hematopoietic stem cells (HSCs) enhances self-renewal and survival. Its expression in specific subtypes of myeloid and lymphoid leukemias provides resistance to apoptosis and blocks further differentiation. A direct oncogenic role in the hematopoietic system has recently been demonstrated by several mouse models. Targets of miR-125b include key proteins regulating apoptosis, innate immunity, inflammation and hematopoietic differentiation. Leukemia (2012) 26, 2011–2018; doi:10.1038/leu.2012.90 Keywords: microRNA; hematopoiesis; hematological malignancies; acute myeloid leukemia; acute lymphoblastic leukemia MicroRNAs (miRNAs) are 21–23-nucleotide non-coding RNAs that nucleotides with the seed region of miR-125b (ebv-miR-BART21-5p, have crucial roles in fundamental biological processes by ebv-miR-BART8 and rlcv-miR-rL1-25). In humans, as in most of the regulating the levels of multiple proteins. They are transcribed genomes, there are two paralogs (hsa-miR-125b-1 on chromosome as primary miRNAs and processed in the nucleus by the RNase III 11 and hsa-miR-125b-2 on chromosome 21), coding for the same endonuclease DROSHA to liberate 70-nucleotide stem loops, the mature sequence. -

Identification of Sequence Variants
Wang et al. BMC Medical Genomics (2019) 12:28 https://doi.org/10.1186/s12920-019-0475-x RESEARCH ARTICLE Open Access Identification of sequence variants associated with severe microtia-astresia by targeted sequencing Pu Wang1, Yibei Wang1, Xinmiao Fan1, Yaping Liu2, Yue Fan1, Tao Liu3, Chongjian Chen3, Shuyang Zhang4*† and Xiaowei Chen1*† Abstract Background: Microtia-atresia is characterized by abnormalities of the auricle (microtia) and aplasia or hypoplasia of the external auditory canal, often associated with middle ear abnormalities. To date, no causal genetic mutations or genes have been identified in microtia-atresia patients. Methods: We designed a panel of 131 genes associated with external/middle or inner ear deformity. Targeted genomic capturing combined with next-generation sequencing (NGS) was utilized to screen for mutations in 40 severe microtia-atresia patients. Mutations detected by NGS were filtered and validated. And then mutations were divided into three categories—rare or novel variants, low-frequency variants and common variants—based on their frequency in the public database. The rare or novel mutations were prioritized by pathogenicity analysis. For the low-frequency variants and common variants, we used association studies to explore risk factors of severe microtia-atresia. Results: Sixty-five rare heterozygous mutations of 42 genes were identified in 27 (67.5%) severe microtia-atresia patients. Association studies to determine genes that were potentially pathogenic found that PLEC, USH2A, FREM2, DCHS1, GLI3, POMT1 and GBA genes were significantly associated with severe microtia-atresia. Of these, DCHS1 was strongly suggested to cause severe microtia-atresia as it was identified by both low-frequency and common variants association studies. -

Specific Aim # 2A to Determine If Mir-122 Down-Regulates OCLN In
NOVEL ROLES OF MICRORNAS IN HEPATITIS C by Hossein Sendi A dissertation submitted to the faculty of The University of North Carolina at Charlotte in partial fulfillment of the requirements for the degree of Doctor of Philosophy in Biology Charlotte 2013 Approved by: ______________________________ Dr. Mark G. Clemens ______________________________ Dr. Herbert L. Bonkovsky ______________________________ Dr. Laura W. Schrum _____________________________ Dr. Valery Grdzelishvili ____________________________ Dr. Kent E. Curran ii ©2013 Hossein Sendi ALL RIGHTS RESERVED iii ABSTRACT HOSSEIN SENDI. Novel roles of microRNAs in hepatitis C. (Under the direction of MARK CLEMENS and HERBERT BONKOVSKY) Approximately, 170 million people are infected by hepatitis C virus (HCV) worldwide, and of these, nearly 85% will develop chronic hepatitis C (CHC). Despite finding new anti-viral treatment that increase response rate from 45 % to 65-70 %, investigations continue to find more effective treatments for hepatitis C because of side effects and limitations of current treatment. It is known that miR-122 enhances HCV replication by binding to two closely spaced target sites in the 5’-UTR of the viral genome, which leads to an increase in abundance of HCV RNA. We found that miR-122 down- regulates Occludin (OCLN), one of the key HCV receptors, by directly targeting 3’-UTR of OCLN mRNA. We also found that interaction of miR-122 with 3’-UTR of OCLN mRNA eventually results in a decrease in HCV entry. In accordance with our in vitro study, we found an inverse correlation between pre-treatment levels of miR-122 and HCV RNA levels in patients with CHC. This is a new finding of our study which is consonant with our hypothesis that miR-122 may play an antiviral role in uninfected hepatocytes and early stages of HCV infection. -

A Catalogue of Stress Granules' Components
Catarina Rodrigues Nunes A Catalogue of Stress Granules’ Components: Implications for Neurodegeneration UNIVERSIDADE DO ALGARVE Departamento de Ciências Biomédicas e Medicina 2019 Catarina Rodrigues Nunes A Catalogue of Stress Granules’ Components: Implications for Neurodegeneration Master in Oncobiology – Molecular Mechanisms of Cancer This work was done under the supervision of: Clévio Nóbrega, Ph.D UNIVERSIDADE DO ALGARVE Departamento de Ciências Biomédicas e Medicina 2019 i ii A catalogue of Stress Granules’ Components: Implications for neurodegeneration Declaração de autoria de trabalho Declaro ser a autora deste trabalho, que é original e inédito. Autores e trabalhos consultados estão devidamente citados no texto e constam na listagem de referências incluída. I declare that I am the author of this work, that is original and unpublished. Authors and works consulted are properly cited in the text and included in the list of references. _______________________________ (Catarina Nunes) iii Copyright © 2019 Catarina Nunes A Universidade do Algarve reserva para si o direito, em conformidade com o disposto no Código do Direito de Autor e dos Direitos Conexos, de arquivar, reproduzir e publicar a obra, independentemente do meio utilizado, bem como de a divulgar através de repositórios científicos e de admitir a sua cópia e distribuição para fins meramente educacionais ou de investigação e não comerciais, conquanto seja dado o devido crédito ao autor e editor respetivos. iv Part of the results of this thesis were published in Nunes,C.; Mestre,I.; Marcelo,A. et al. MSGP: the first database of the protein components of the mammalian stress granules. Database (2019) Vol. 2019. (In annex A). v vi ACKNOWLEDGEMENTS A realização desta tese marca o final de uma etapa académica muito especial e que jamais irei esquecer. -

Novel Targets of Apparently Idiopathic Male Infertility
International Journal of Molecular Sciences Review Molecular Biology of Spermatogenesis: Novel Targets of Apparently Idiopathic Male Infertility Rossella Cannarella * , Rosita A. Condorelli , Laura M. Mongioì, Sandro La Vignera * and Aldo E. Calogero Department of Clinical and Experimental Medicine, University of Catania, 95123 Catania, Italy; [email protected] (R.A.C.); [email protected] (L.M.M.); [email protected] (A.E.C.) * Correspondence: [email protected] (R.C.); [email protected] (S.L.V.) Received: 8 February 2020; Accepted: 2 March 2020; Published: 3 March 2020 Abstract: Male infertility affects half of infertile couples and, currently, a relevant percentage of cases of male infertility is considered as idiopathic. Although the male contribution to human fertilization has traditionally been restricted to sperm DNA, current evidence suggest that a relevant number of sperm transcripts and proteins are involved in acrosome reactions, sperm-oocyte fusion and, once released into the oocyte, embryo growth and development. The aim of this review is to provide updated and comprehensive insight into the molecular biology of spermatogenesis, including evidence on spermatogenetic failure and underlining the role of the sperm-carried molecular factors involved in oocyte fertilization and embryo growth. This represents the first step in the identification of new possible diagnostic and, possibly, therapeutic markers in the field of apparently idiopathic male infertility. Keywords: spermatogenetic failure; embryo growth; male infertility; spermatogenesis; recurrent pregnancy loss; sperm proteome; DNA fragmentation; sperm transcriptome 1. Introduction Infertility is a widespread condition in industrialized countries, affecting up to 15% of couples of childbearing age [1]. It is defined as the inability to achieve conception after 1–2 years of unprotected sexual intercourse [2]. -

Interferon-Inducible Antiviral Effectors
REVIEWS Interferon-inducible antiviral effectors Anthony J. Sadler and Bryan R. G. Williams Abstract | Since the discovery of interferons (IFNs), considerable progress has been made in describing the nature of the cytokines themselves, the signalling components that direct the cell response and their antiviral activities. Gene targeting studies have distinguished four main effector pathways of the IFN-mediated antiviral response: the Mx GTPase pathway, the 2′,5′-oligoadenylate-synthetase-directed ribonuclease L pathway, the protein kinase R pathway and the ISG15 ubiquitin-like pathway. As discussed in this Review, these effector pathways individually block viral transcription, degrade viral RNA, inhibit translation and modify protein function to control all steps of viral replication. Ongoing research continues to expose additional activities for these effector proteins and has revealed unanticipated functions of the antiviral response. Pattern-recognition Interferon (IFN) was discovered more than 50 years ago in components of the IFNR signalling pathway (STAT1 receptors as an agent that inhibited the replication of influenza (signal transducer and activator of transcription 1), TYK2 (PRRs). Host receptors that can virus1. The IFN family of cytokines is now recognized as (tyrosine kinase 2) or UNC93B) die of viral disease, with sense pathogen-associated a key component of the innate immune response and the the defect in IFNAR (rather than IFNGR) signalling molecular patterns and initiate 6–9 signalling cascades that lead to first line of defence against viral infection. Accordingly, having the more significant role . an innate immune response. IFNs are currently used therapeutically, with the most The binding of type I IFNs to the IFNAR initiates a These can be membrane bound noteworthy example being the treatment of hepatitis C signalling cascade, which leads to the induction of more (such as Toll-like receptors) or virus (HCV) infection, and they are also used against than 300 IFN-stimulated genes (ISGs)10. -

125B Occurs Through Evolving Mirna–Target Gene Pairs
Conserved Regulation of p53 Network Dosage by MicroRNA– 125b Occurs through Evolving miRNA–Target Gene Pairs The MIT Faculty has made this article openly available. Please share how this access benefits you. Your story matters. Citation Le, Minh T. N. et al. “Conserved Regulation of P53 Network Dosage by MicroRNA–125b Occurs Through Evolving miRNA–Target Gene Pairs.” Ed. Michael T. McManus. PLoS Genetics 7.9 (2011): e1002242. Web. 10 Feb. 2012. As Published http://dx.doi.org/10.1371/journal.pgen.1002242 Publisher Public Library of Science Version Final published version Citable link http://hdl.handle.net/1721.1/69088 Terms of Use Creative Commons Attribution Detailed Terms http://creativecommons.org/licenses/by/2.5/ Conserved Regulation of p53 Network Dosage by MicroRNA–125b Occurs through Evolving miRNA–Target Gene Pairs Minh T. N. Le1,2,3., Ng Shyh-Chang1,4., Swea Ling Khaw1,5, Lingzi Chin1, Cathleen Teh6, Junliang Tay1, Elizabeth O’Day2, Vladimir Korzh5, Henry Yang7, Ashish Lal2,8, Judy Lieberman2, Harvey F. Lodish3,9,10*, Bing Lim1,3,11* 1 Stem Cell and Developmental Biology, Genome Institute of Singapore, Singapore, Singapore, 2 Immune Disease Institute and Program in Cellular and Molecular Medicine, Children’s Hospital Boston, Harvard Medical School, Boston, Massachusetts, United States of America, 3 Computation and Systems Biology, Singapore–MIT Alliance, Singapore, Singapore, 4 Department of Biological Chemistry and Molecular Pharmacology, Harvard Medical School, Boston, Massachusetts, United States of America, 5 NUS Graduate -

Missense Mutation in the Second RNA Binding Domain Reveals a Role for Prkra (PACT/RAX) During Skull Development Benjamin K
Missense mutation in the second RNA binding domain reveals a role for Prkra (PACT/RAX) during skull development Benjamin K. Dickerman, Christine L. White, Claire Chevalier, Valerie Nalesso, Cyril Charles, Sophie Fouchécourt, Florian Jean Louis Guillou, Laurent Viriot, Ganes C. Sen, Yann Hérault To cite this version: Benjamin K. Dickerman, Christine L. White, Claire Chevalier, Valerie Nalesso, Cyril Charles, et al.. Missense mutation in the second RNA binding domain reveals a role for Prkra (PACT/RAX) during skull development. PLoS ONE, Public Library of Science, 2011, 6 (12), pp.e28537. 10.1371/jour- nal.pone.0028537. hal-01129622 HAL Id: hal-01129622 https://hal.archives-ouvertes.fr/hal-01129622 Submitted on 29 May 2020 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. Missense Mutation in the Second RNA Binding Domain Reveals a Role for Prkra (PACT/RAX) during Skull Development Benjamin K. Dickerman1,2, Christine L. White1, Claire Chevalier3, Vale´rie Nalesso3, Cyril Charles4, Sophie Fouche´court5, Florian Guillou5, Laurent Viriot4, Ganes C. Sen1,2*, -

The Human GCOM1 Complex Gene Interacts with the NMDA Receptor and Internexin-Alpha☆
HHS Public Access Author manuscript Author ManuscriptAuthor Manuscript Author Gene. Author Manuscript Author manuscript; Manuscript Author available in PMC 2018 June 20. Published in final edited form as: Gene. 2018 March 30; 648: 42–53. doi:10.1016/j.gene.2018.01.029. The human GCOM1 complex gene interacts with the NMDA receptor and internexin-alpha☆ Raymond S. Roginskia,b,*, Chi W. Laua,1, Phillip P. Santoiemmab,2, Sara J. Weaverc,3, Peicheng Dud, Patricia Soteropoulose, and Jay Yangc aDepartment of Anesthesiology, CMC VA Medical Center, Philadelphia, PA 19104, United States bDepartment of Anesthesiology and Critical Care, Perelman School of Medicine, University of Pennsylvania, Philadelphia, PA 19104, United States cDepartment of Anesthesia, University of Wisconsin at Madison, Madison, WI 53706, United States dBioinformatics, Rutgers-New Jersey Medical School, Newark, NJ 07103, United States eDepartment of Genetics, Rutgers-New Jersey Medical School, Newark, NJ 07103, United States Abstract The known functions of the human GCOM1 complex hub gene include transcription elongation and the intercalated disk of cardiac myocytes. However, in all likelihood, the gene's most interesting, and thus far least understood, roles will be found in the central nervous system. To investigate the functions of the GCOM1 gene in the CNS, we have cloned human and rat brain cDNAs encoding novel, 105 kDa GCOM1 combined (Gcom) proteins, designated Gcom15, and identified a new group of GCOM1 interacting genes, termed Gints, from yeast two-hybrid (Y2H) screens. We showed that Gcom15 interacts with the NR1 subunit of the NMDA receptor by co- expression in heterologous cells, in which we observed bi-directional co-immunoprecipitation of human Gcom15 and murine NR1. -

Mutation Discovery in Mice by Whole Exome Sequencing
Fairfield et al. Genome Biology 2011, 12:R86 http://genomebiology.com/2011/12/9/R86 METHOD Open Access Mutation discovery in mice by whole exome sequencing Heather Fairfield1, Griffith J Gilbert1, Mary Barter1, Rebecca R Corrigan2, Michelle Curtain1, Yueming Ding3, Mark D’Ascenzo4, Daniel J Gerhardt4, Chao He5, Wenhui Huang6, Todd Richmond4, Lucy Rowe1, Frank J Probst2, David E Bergstrom1, Stephen A Murray1, Carol Bult1, Joel Richardson1, Benjamin T Kile7, Ivo Gut8, Jorg Hager8, Snaevar Sigurdsson9, Evan Mauceli9, Federica Di Palma9, Kerstin Lindblad-Toh9, Michael L Cunningham10, Timothy C Cox10, Monica J Justice2, Mona S Spector5, Scott W Lowe5, Thomas Albert4, Leah Rae Donahue1, Jeffrey Jeddeloh4, Jay Shendure10 and Laura G Reinholdt1* Abstract We report the development and optimization of reagents for in-solution, hybridization-based capture of the mouse exome. By validating this approach in a multiple inbred strains and in novel mutant strains, we show that whole exome sequencing is a robust approach for discovery of putative mutations, irrespective of strain background. We found strong candidate mutations for the majority of mutant exomes sequenced, including new models of orofacial clefting, urogenital dysmorphology, kyphosis and autoimmune hepatitis. Background burdensome and expensive for many laboratories. Targeted Phenotype-driven approaches in model organisms, includ- sequencing approaches are less expensive and the data are ing spontaneous mutation discovery, standard N-ethyl-N- accordingly more manageable, but this technique requires nitrosourea (ENU) mutagenesis screens, sensitized screens substantial genetic mapping and the design and purchase and modifier screens, are established approaches in func- of custom capture tools (that is, arrays or probe pools) [4]. -

Host Protein Interactions
scientificscientificreport report Structure homology and interaction redundancy for discovering virus–host protein interactions Benoıˆt de Chassey 1,Laure`ne Meyniel-Schicklin 2,3, Anne Aublin-Gex 2,3, Vincent Navratil 2,3,w, Thibaut Chantier 2,3, Patrice Andre´1,2,3 &VincentLotteau2,3+ 1Hospices Civils de Lyon, Hoˆpital de la Croix-Rousse, Laboratoire de virologie, Lyon , 2Inserm, U1111, Lyon , and 3Universite´ de Lyon, Lyon, France Virus–host interactomes are instrumental to understand global purification coupled to mass spectrometry, led to the publication perturbations of cellular functions induced by infection and of the first virus–host interactomes [3–4]. Although incorporation discover new therapies. The construction of such interactomes is, of data from different methods or variation of the same method [5] however, technically challenging and time consuming. Here we has improved the quality of the data sets, diversification of describe an original method for the prediction of high-confidence methods is still clearly needed to generate high-quality interactions between viral and human proteins through comprehensive virus–host interactomes. In addition, regarding a combination of structure and high-quality interactome data. the size of host genome and the huge diversity of viruses, millions Validation was performed for the NS1 protein of the influenza of binary interactions remain to be tested. Therefore, accurate and virus, which led to the identification of new host factors that rapid methods for the identification of cellular interactors control viral replication. controlling viral replication is a major issue and a crucial step Keywords: interactome; prediction; protein interaction; towards the selection of original therapeutic targets and drug structure; virus development. -

LGP2 Binds to PACT to Regulate RIG-I– and MDA5-Mediated Antiviral
LGP2 binds to PACT to regulate RIG-I– and MDA5-mediated antiviral responses Raul Sanchez David, Chantal Combredet, Valérie Najburg, Gaël Millot, Guillaume Beauclair, Benno Schwikowski, Thibaut Léger, Jean-Michel Camadro, Yves Jacob, Jacques Bellalou, et al. To cite this version: Raul Sanchez David, Chantal Combredet, Valérie Najburg, Gaël Millot, Guillaume Beauclair, et al.. LGP2 binds to PACT to regulate RIG-I– and MDA5-mediated antiviral responses. Science Signaling, American Association for the Advancement of Science, 2019, 12 (601), pp.eaar3993. 10.1126/scisig- nal.aar3993. hal-03100138 HAL Id: hal-03100138 https://hal-cnrs.archives-ouvertes.fr/hal-03100138 Submitted on 6 Jan 2021 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. SCIENCE SIGNALING | RESEARCH ARTICLE IMMUNOLOGY Copyright © 2019 The Authors, some rights reserved; LGP2 binds to PACT to regulate RIG-I– and exclusive licensee American Association MDA5-mediated antiviral responses for the Advancement Raul Y. Sanchez David1,2*†, Chantal Combredet1*, Valérie Najburg1, Gael A. Millot3, of Science. No claim 1 4 5 5,6 to original U.S. Guillaume Beauclair , Benno Schwikowski , Thibaut Léger , Jean-Michel Camadro , Government Works Yves Jacob7, Jacques Bellalou8, Nolwenn Jouvenet1, Frédéric Tangy1‡, Anastassia V.