Schwann Cell Precursors Generate the Majority of Chromaffin Cells in Zuckerkandl Organ and Some Sympathetic Neurons in Paragangl
Total Page:16
File Type:pdf, Size:1020Kb

Load more
Recommended publications
-

Expression Pattern of Delta-Like 1 Homolog in Developing Sympathetic Neurons and Chromaffin Cells
Published in "Gene Expression Patterns 30: 49–54, 2018" which should be cited to refer to this work. Expression pattern of delta-like 1 homolog in developing sympathetic neurons and chromaffin cells ∗ Tehani El Faitwria,b, Katrin Hubera,c, a Institute of Anatomy & Cell Biology, Albert-Ludwigs-University Freiburg, Albert-Str. 17, 79104, Freiburg, Germany b Department of Histology and Anatomy, Faculty of Medicine, Benghazi University, Benghazi, Libya c Department of Medicine, University of Fribourg, Route Albert-Gockel 1, 1700, Fribourg, Switzerland ABSTRACT Keywords: Delta-like 1 homolog (DLK1) is a member of the epidermal growth factor (EGF)-like family and an atypical notch Sympathetic neurons ligand that is widely expressed during early mammalian development with putative functions in the regulation Chromaffin cells of cell differentiation and proliferation. During later stages of development, DLK1 is downregulated and becomes DLK1 increasingly restricted to specific cell types, including several types of endocrine cells. DLK1 has been linked to Adrenal gland various tumors and associated with tumor stem cell features. Sympathoadrenal precursors are neural crest de- Organ of Zuckerkandl rived cells that give rise to either sympathetic neurons of the autonomic nervous system or the endocrine Development ffi Neural crest chroma n cells located in the adrenal medulla or extraadrenal positions. As these cells are the putative cellular Phox2B origin of neuroblastoma, one of the most common malignant tumors in early childhood, their molecular char- acterization is of high clinical importance. In this study we have examined the precise spatiotemporal expression of DLK1 in developing sympathoadrenal cells. We show that DLK1 mRNA is highly expressed in early sympa- thetic neuron progenitors and that its expression depends on the presence of Phox2B. -

I. Introduction B. Adrenal Cortex
I. Introduction Adrenal Glands • suprarenal – they sit on top of the kidneys • each is composed of 2 distinct regions: A. Adrenal Medulla - the inner region - comprises 20% of the gland - secretes epinephrine and norepinephrine - derived from ectoderm B. Adrenal Cortex 1) Zona Glomerulosa (outermost region) - produces mineralocorticoids (aldosterone) • the outer region 2) Zona Fasiculata (middle region) - produces glucocorticoids (cortisol) as well as • comprises 80% of the gland estrogens and androgens • secretes corticosteroids 3) Zona Reticularis (innermost region) • derived from mesoderm - same function as zona fasiculata DHEA – dehydroepiandrosterone • an adrenal androgen in females • responsible for growth of pubic and axillary hair C. Pathologies Associated with Adrenal II. Mineralocorticoids (Aldosterone) Androgen Hypersecretion A. Functions 1.Adrenogenital Syndrome - promotes reabsorption of Na+ and - hypersecretion of androgens or estrogens secretion of K+ from the distal portion of the a) in the adult female: nephron..primary regulator of salt balance and extracellular volume - masculinization (i.e. hirsutism) -Similar (but less important) effect on salt b) in the female embryo: transport in colon, salivary glands, and - female pseudohermaphroditism sweat glands. c) in the adult male: - no effect d) in young boys: - precocious pseudopuberty 1 II. Mineralocorticoids (Aldosterone) C. Pathologies B. Regulation of Secretion 1. Hypersecretion 1. Renin Angiotensin a. primary hyperaldosteronism - Angiotensin II stimulates aldost. secretion - Conn’s syndrome 2. Potassium - usually due to a tumor on the gland + - high levels of K induce aldost. secretion - too much secretion of gland itself 3. ACTH –no direct role b. secondary hyperaldosteronism - default in renin angiotensin system - most common in atherosclerosis of renal arteries C. Pathologies III. Glucocorticoids (Cortisol) 1. -
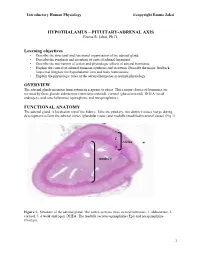
HYPOTHALAMUS – PITUITARY-ADRENAL AXIS Learning Objectives OVERVIEW FUNCTIONAL ANATOMY
Introductory Human Physiology ©copyright Emma Jakoi HYPOTHALAMUS – PITUITARY-ADRENAL AXIS Emma R. Jakoi, Ph.D. Learning objectives • Describe the structural and functional organization of the adrenal gland. • Describe the synthesis and secretion of cortical adrenal hormones. • Describe the mechanism of action and physiologic effects of adrenal hormones. • Explain the control of adrenal hormone synthesis and secretion. Describe the major feedback loops that integrate the hypothalamic axis and body homeostasis. • Explain the physiologic roles of the adrenal hormones in normal physiology. OVERVIEW The adrenal glands maintain homeostasis in response to stress. Three major classes of hormones are secreted by these glands: aldosterone (mineralocorticoid), cortisol (glucocorticoid), DHEA (weak androgen), and catecholamines (epinephrine and norepinephrine). FUNCTIONAL ANATOMY The adrenal gland is located on top of the kidney. Like the pituitary, two distinct tissues merge during development to form the adrenal cortex (glandular tissue) and medulla (modified neuronal tissue) (Fig 1). 1 2 cortex 3 medulla Figure 1. Structure of the adrenal gland. The cortex secretes three steroid hormones: 1. aldosterone, 2. cortisol, 3. a weak androgen, DHEA. The medulla secretes epinephrine (Epi) and norepinephrine (NorEpi). 1 Introductory Human Physiology ©copyright Emma Jakoi MINERALOCORTICOIDS The major mineralocorticoid in humans is aldosterone. Aldosterone is NOT under the hypothalamus- pituitary control and does not mediate a negative feedback to this axis. Aldosterone secretion is increased by the vasoconstrictor, angiotensin II, and by elevated plasma K+ concentration. Elevated plasma Na+ inhibits the secretion of aldosterone. Aldosterone, acts in the kidney to promote secretion of K+ into the urine from the blood and the reabsorption of Na+ from the urine into the blood. -

Novel Autocrine Feedback Control of Catecholamine Release a Discrete Chromogranin a Fragment Is a Noncompetitive Nicotinic Cholinergic Antagonist Sushil K
Novel Autocrine Feedback Control of Catecholamine Release A Discrete Chromogranin A Fragment is a Noncompetitive Nicotinic Cholinergic Antagonist Sushil K. Mahata,* Daniel T. O’Connor,* Manjula Mahata,* Seung Hyun Yoo,‡ Laurent Taupenot,* Hongjiang Wu,* Bruce M. Gill,* and Robert J. Parmer* *Department of Medicine and Center for Molecular Genetics, University of California, San Diego, California 92093 and Department of Veterans Affairs Medical Center, San Diego, California 92161, and ‡National Institute of Deafness and Communicative Disorders, National Institutes of Health, Bethesda, Maryland 20892 Abstract completely established, recent evidence implicates chromogra- nin A as a precursor of several small biologically active secre- Catecholamine secretory vesicle core proteins (chromog- tion-inhibitory peptides that may play an autocrine regulatory ranins) contain an activity that inhibits catecholamine re- role in neuroendocrine secretion from a variety of cell types (3, lease, but the identity of the responsible peptide has been 7–10). The primary structure of chromogranin A reveals 8–10 elusive. Size-fractionated chromogranins antagonized nico- conserved pairs of basic amino acids that represent potential tinic cholinergic-stimulated catecholamine secretion; the in- proteolytic cleavage sites for the generation of such peptides hibitor was enriched in processed chromogranin fragments, (11–17), and chromogranin A is a substrate for the prohor- and was liberated from purified chromogranin A. Of 15 syn- mone convertases (18–20). of chromogranin A, one To date, several chromogranin A–derived peptides have %80 ف thetic peptides spanning (bovine chromogranin A344–364 [RSMRLSFRARGYGFRG- been identified that affect the secretory function of the parent PGLQL], or catestatin) was a potent, dose-dependent (IC50 cells. -

Dream Recall/Affect and the Hypothalamic–Pituitary–Adrenal Axis
Review Dream Recall/Affect and the Hypothalamic–Pituitary–Adrenal Axis Athanasios Tselebis 1, Emmanouil Zoumakis 2 and Ioannis Ilias 3,* 1 Department of Psychiatry, Sotiria Hospital, 115 27 Athens, Greece; [email protected] 2 First Department of Pediatrics, National and Kapodistrian University of Athens Medical School, Aghia Sophia Children’s Hospital, 115 27 Athens, Greece; [email protected] 3 Department of Endocrinology, Diabetes and Metabolism, Elena Venizelou Hospital, 115 21 Athens, Greece * Correspondence: [email protected]; Tel.: +30-213-205-1389 Abstract: In this concise review, we present an overview of research on dream recall/affect and of the hypothalamic–pituitary–adrenal (HPA) axis, discussing caveats regarding the action of hormones of the HPA axis (mainly cortisol and its free form, cortisol-binding globulin and glucocorticoid receptors). We present results of studies regarding dream recall/affect and the HPA axis under physiological (such as waking) or pathological conditions (such as in Cushing’s syndrome or stressful situations). Finally, we try to integrate the effect of the current COVID-19 situation with dream recall/affect vis-à-vis the HPA axis. Keywords: dreams; cortisol; stress; memory; sleep; HPA axis 1. Introduction—Dream Recall Almost all humans dream (indeed, there may be 0.5% of people who do not do so) [1]. Dreams are a series of images, thoughts and senses and are considered a specific type Citation: Tselebis, A.; Zoumakis, E.; of experience that occurs in our brain during sleep. During the dream, there is limited Ilias, I. Dream Recall/Affect and the control of the dream content, visual images and memory activation. -

Pig Gonads, Adrenal Glands and Brain C
Immunoreactive cytochrome P-45017\g=a\in rat and guinea- pig gonads, adrenal glands and brain C. Le Goascogne1, N. Sanan\l=e'\s1, M. Gou\l=e'\zou1, S. Takemori2, S. Kominami2, E. E. Baulieu1 and P. Robel1 1INSERM U33, Communications Hormonales, and Faculté de Médecine, Université Paris-sud, Lab Hormones F-94275 Bicêtre Cedex France 2 Faculty of Integrated Arts and Sciences, Hiroshima University, Hiroshima 730, Japan Summary. The cytochrome P-45017\g=a\-hydroxylase, 17\ar=r\20lyase (P-45017\g=a\) is the key enzyme responsible for the biosynthesis of androgens in steroidogenic organs. Its cellular localization has been examined with an immunohistochemical technique. In immature rat ovary, P-45017\g=a\was first detected in sparse interstitial cells on postnatal Day 8. The number of immunoreactive interstitial cells increased thereafter and the intensity of P-45017\g=a\staining in these cells was highest at 3 weeks of age. The intensity of staining then started to decline and was very faint at Day 35. From 6 weeks on, the distribution of immunoreactive P-45017\g=a\was of the adult type: it was detected exclusively in the thecal cells of the large antral, preovulatory, follicles. P-45017\g=a\was not detectable during pregnancy except on the day of parturition, when thecal cells were transiently immunoreactive. The staining had vanished 24 h after delivery. Human chorionic gonadotrophin (hCG), injected into immature females on Days 24 to 26, induced P-45017\g=a\prematurely in thecal cells. When injected on Days 12 to 14 of pregnancy, hCG also induced P-45017\g=a\in the thecal cells surrounding the largest follicles, whereas the interstitial and luteal cells were not immunostained. -

Extra-Adrenal Chromaffin Cells of the Zuckerkandl´S Paraganglion: Morphological and Electrophysiological Study
275 Extra-adrenal chromaffin cells of the Zuckerkandl´s paraganglion: morphological and electrophysiological study. Beatriz Galán-Rodríguez, M. Pilar Ramírez-Ponce, Fadwa El Banoua, Juan A. Flores, Juan Bellido and Emilio Fernández-Espejo. Departamento de Fisiología Médica y Biofísica. Universidad de Sevilla. Spain. Correspondence: Dra. Beatriz Galán Rodríguez or Dr. Emilio Fernández-Espejo, Departamento de Fisiología Médica y Biofísica, Facultad de Medicina, Universidad de Sevilla, 41009. Sevilla. Spain. Phone: 34-954-556584; Fax: 34-954-551769; Email: [email protected] ; [email protected] Cell Biology of the Chromaffin Cell R. Borges & L. Gandía Eds. Instituto Teófilo Hernando, Spain, 2004 Cell Biology of the Chromaffin Cell 276 Parkinson´s disease is one of the most important neurodegenerative disorders that affects to one out of a hundred of the world population elder than 65. It has been observed in our laboratory, for the first time, that intrabrain transplantation of chromaffin cell aggregates from the Zuckerkandl´s organ, an extraadrenal paraganglion located adjacent to the lower abdominal aorta, induced gradual improvement of functional deficits in animal models of Parkinson´s disease1. This functional regeneration was likely caused by long-survival of grafted cells and chronic trophic action of dopaminotrophic factors, glial cell line-derived 2,3 4,5 factor (GDNF) and transforming growth factor beta1 (TGF-b1) , which are expressed and delivered by long-surviving grafted chromaffin cells. The objective of this study is to discern the morphological and cytological characteristics of extra-adrenal cells of the Zuckerkandl’s organ. On the other hand, long survival of extra-adrenal chromaffin cells could be related to resistance to hypoxia, since it is certainly know that hypoxia is a primary factor involved in cell death after intrabrain grafting. -

Adrenal Gland Hormones
CHAPTER 8 Adrenal Gland Hormones Devra K. Dang, PharmD, BCPS, CDE, FNAP | Trinh Pham, PharmD, BCOP | Jennifer J. Lee, PharmD, BCPS, CDE LEARNING OBJECTIVES KEY TERMS AND DEFINITIONS After completing this chapter, you should be able to ACTH (adrenocorticotropic hormone) — a hormone produced 1. Identify the hormones produced by the adrenal glands by the pituitary gland that stimulates 2. Describe the functions of mineralocorticoids and glucocorticoids in the body the adrenal cortex to produce glucocorticoids, mineralocorticoids, 3. Recognize the signs and symptoms of adrenal insuffi ciency and androgens. PART 4. Describe the pharmacological treatment of patients with acute and chronic adrenal Addison ’ s disease — a disorder insuffi ciency in which the adrenal glands do not produce enough steroid hormones. 3 5. Recognize the signs and symptoms of Cushing ’ s syndrome and the result of too Adenoma — a benign much cortisol (noncancerous) tumor of glandular 6. Describe the pharmacologic and nonpharmacologic management of patients with origin. Cushing ’ s syndrome Adrenal insuffi ciency — a term 7. List management strategies for administration of glucocorticoid and mineralocorti- referring to a defi ciency in the levels of adrenal hormones. coid therapy to avoid development of adrenal disorders Aldosterone — the hormone produced by the adrenal glands that regulates the balance of sodium, he adrenal glands are an integral part of the endocrine system, secreting water, and potassium concentrations in the body. T hormones that act throughout the body to regulate functions and promote Corticotropin-releasing homeostasis. In addition to the neurotransmitters epinephrine and norepineph- hormone (CRH) — a hormone rine, the corticosteroids secreted by the adrenal glands are vital to a wide released by the hypothalamus that variety of physiological processes. -

Cell Proliferation
Proc. Nati. Acad. Sci. USA Vol. 91, pp. 1771-1775, March 1994 Neurobiology Insulin-like growth factors act synergistically with basic fibroblast growth factor and nerve growth factor to promote chromaffin cell proliferation MORTEN FRODIN* AND STEEN GAMMELTOFT*t Department of Clinical Chemistry, Bispebjerg Hospital, DK 2400 Copenhagen NV, Denmark Communicated by Viktor Mutt, November 29, 1993 (receivedfor review August 26, 1993) ABSTRACT We have investigated the effects of insulin- addition, NGF and bFGF stimulate transdifferentiation of like growth factors (IGFs), basic fibroblast growth factor cultured chromaffin cells into a sympathetic neuron-like (bFGF), and nerve growth factor (NGF) on DNA synthesis in phenotype and increase cell survival and catecholamine cultured chromaffin cells from fetal, neonatal, and adult rats synthesis (8, 9, 13-15). In adult rats, chromaffmi-cell replace- by using 5-bromo-2'-deoxyuridine (BrdUrd) pulse labeling for ment decreases after denervation of the adrenal medulla, 24 or 48 h and immunocytochemical staining of cell nuclei. suggesting that neurogenic stimuli can regulate chromaffmi- After 6 days in culture in the absence ofgrowth factors, nuclear cell proliferation (16). BrdUrd incorporation was detected in 30% offetal chromafFin Insulin-like growth factor I and II (IGF-I and IGF-II, cells, 1.5% of neonatal cells, and 0.1% of adult cells. Addition respectively) act as neurotrophic factors in various parts of of 10 nM IGF-I or IGF-ll increased the fraction of BrdUrd- the developing and adult nervous system (17). IGF-II mRNA labeled nuclei to 50% offetal, 20% ofneonatal, and 2% ofadult and immunoreactivity have been demonstrated in adult hu- chromaffn cells. -

The Generation of Monoclonal Antibodies That Bind Preferentially to Adrenal Chromaffin Cells and the Cells of Embryonic Sympathetic Ganglia
The Journal of Neuroscience, November 1991, 7 7(11): 34933506 The Generation of Monoclonal Antibodies that Bind Preferentially to Adrenal Chromaffin Cells and the Cells of Embryonic Sympathetic Ganglia Josette F. Carnahaw and Paul l-i. Patterson Biology Division, California Institute of Technology, Pasadena, California 91125 Adrenal chromaffin ceils, sympathetic neurons, and small technical limitation in this effort is the lack of markers specific intensely fluorescent (SIF) cells are each derived from the for the various stagesof development within a particular lineage. neural crest, produce catecholamines, and share certain Without such markers, cells making key phenotypic decisions morphological features. These cell types are also partially cannot be identified. It has been especially difficult to find mo- interconvertible in cell culture (Doupe et al., 1985a,b; An- lecular labels for committed progenitor cells. Monoclonal an- derson and Axel, 1988). Thus, these cells are said to be tibodies can be used as highly specific markers, and the hybrid- members of the sympathoadrenal (SA) lineage and could oma method can be manipulated so asto enhancethe likelihood share a common progenitor. To investigate the origins of this of obtaining antibodies against rare antigens of interest (Mat- lineage further, we used the cyclophosphamide immuno- thew and Patterson, 1983; Barald and Wessels,1984; Agius and suppression method (Matthew and Patterson, 1983) to gen- Richman, 1986; Barclay and Smith, 1986; Golumbeski and Di- erate five monoclonal antibodies (SAl-5) that bind strongly mond, 1986; Hockfield, 1987; Mahana et al., 1987; Matthew to chromaffin cells, with little or no labeling of sympathetic and Sandrock, 1987; Norton and Benjamini, 1987; Huse et al., neurons or SIF cells in frozen sections from adult rats. -

Adrenal and Paraganglia Tumors Adrenal Cortical Tumors IHC: (+) SF1, Inhibin, Melan-A, Calretinin, Synaptophysin, (-) Chromogranin, Cytokeratin, S100
Last updated: 11/11/2020 Prepared by Kurt Schaberg Adrenal and Paraganglia Tumors Adrenal Cortical Tumors IHC: (+) SF1, Inhibin, Melan-A, Calretinin, Synaptophysin, (-) Chromogranin, Cytokeratin, S100. Often variable!! Adrenal Cortical Adenoma Benign. Very common. Often incidentally identified. Usually unilateral solitary masses with atrophic background adrenal gland. Tumor cells can be lipid-rich (clearer) or lipid-poor (pinker) arranged in nests and cords separated by abundant vasculature. Occasional lipofuscin pigment. Nuclei generally small and round (occasional extreme “endocrine atypia” is common). Low/no mitotic activity. Intact reticulin framework. On a spectrum with and may hard to differentiate from hyperplastic nodules, which is more often multinodular (background hyperplasia) and bilateral. Can be non-functional (85%) or functional (15%). Associated with MEN1, FAP, Carney Complex, among Aldosterone-producing→ “Conn syndrome”→ others… hypertension and hypokalemia If aldosterone-secreting adenoma is treated with Cortisol-producing→ (ACTH-independent) spironolactone→ “spironolactone bodies” (below) “Cushing Syndrome” → central obesity, moon face, hirsutism, poor healing, striae Sex-hormone-producing→ Rare (more common in carcinomas). Symptoms depend on hormone/sex (virilization or feminization) Adrenal Cortical Carcinoma Malignant. Most common in older adults. Can present with an incidental unilateral mass or with an endocrinopathy (see above). Solid, broad trabeculae, or large nested growth (more diffuse, and larger groups than in adenomas) Thick fibrous capsule with occasional fibrous bands. Frequent tumor necrosis. Frequent vascular or capsular invasion. Increased mitotic activity. Variants: Oncocytic, Myxoid, Sarcomatoid Mostly sporadic, but can be associated with Lynch Syndrome and Li-Fraumeni Syndrome Distinguishing between an Adrenal Cortical Adenoma vs Carcinoma Weiss Criteria: Weiss Criteria (≥3 = Malignant) Most widely used system, but doesn’t work as well High nuclear grade (based of Fuhrman criteria) in borderline cases or variants. -

Hypothalamic–Pituitary–Adrenal Axis Dysregulation and Behavioral Analysis of Mouse Mutants with Altered Glucocorticoid Or Mineralocorticoid Receptor Function
Stress, September 2008; 11(5): 321–338 REVIEW Hypothalamic–pituitary–adrenal axis dysregulation and behavioral analysis of mouse mutants with altered glucocorticoid or mineralocorticoid receptor function BENEDICT J. KOLBER, LINDSAY WIECZOREK, & LOUIS J. MUGLIA Departments of Pediatrics and Molecular Biology and Pharmacology and Program in Neuroscience, Washington University in St Louis, St Louis, MO 63110, USA (Received 12 July 2007; revised 29 October 2007; accepted 19 November 2007) Abstract Corticosteroid receptors are critical for the maintenance of homeostasis after both psychological and physiological stress. To understand the different roles and interactions of the glucocorticoid receptor (GR) and mineralocorticoid receptor (MR) during stress, it is necessary to dissect the role of corticosteroid signaling at both the system and sub-system level. A variety of GR transgenic mouse lines have recently been used to characterize the role of GR in the CNS as a whole and particularly in the forebrain. We will describe both the behavioral and cellular/molecular implications of disrupting GR function in these animal models and describe the implications of this data for our understanding of normal endocrine function and stress adaptation. MRs in tight epithelia have a long established role in sodium homeostasis. Recently however, evidence has suggested that MRs in the limbic brain also play an important role in psychological stress. Just as with GR, targeted mutations in MR induce a variety of behavioral changes associated with stress adaptation. In this review, we will discuss the implications of this work on MR. Finally, we will discuss the possible interaction between MR and GR and how future work using double mutants (through For personal use only.