Hypothalamic–Pituitary–Adrenal Axis Dysregulation and Behavioral Analysis of Mouse Mutants with Altered Glucocorticoid Or Mineralocorticoid Receptor Function
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Expression Pattern of Delta-Like 1 Homolog in Developing Sympathetic Neurons and Chromaffin Cells
Published in "Gene Expression Patterns 30: 49–54, 2018" which should be cited to refer to this work. Expression pattern of delta-like 1 homolog in developing sympathetic neurons and chromaffin cells ∗ Tehani El Faitwria,b, Katrin Hubera,c, a Institute of Anatomy & Cell Biology, Albert-Ludwigs-University Freiburg, Albert-Str. 17, 79104, Freiburg, Germany b Department of Histology and Anatomy, Faculty of Medicine, Benghazi University, Benghazi, Libya c Department of Medicine, University of Fribourg, Route Albert-Gockel 1, 1700, Fribourg, Switzerland ABSTRACT Keywords: Delta-like 1 homolog (DLK1) is a member of the epidermal growth factor (EGF)-like family and an atypical notch Sympathetic neurons ligand that is widely expressed during early mammalian development with putative functions in the regulation Chromaffin cells of cell differentiation and proliferation. During later stages of development, DLK1 is downregulated and becomes DLK1 increasingly restricted to specific cell types, including several types of endocrine cells. DLK1 has been linked to Adrenal gland various tumors and associated with tumor stem cell features. Sympathoadrenal precursors are neural crest de- Organ of Zuckerkandl rived cells that give rise to either sympathetic neurons of the autonomic nervous system or the endocrine Development ffi Neural crest chroma n cells located in the adrenal medulla or extraadrenal positions. As these cells are the putative cellular Phox2B origin of neuroblastoma, one of the most common malignant tumors in early childhood, their molecular char- acterization is of high clinical importance. In this study we have examined the precise spatiotemporal expression of DLK1 in developing sympathoadrenal cells. We show that DLK1 mRNA is highly expressed in early sympa- thetic neuron progenitors and that its expression depends on the presence of Phox2B. -

I. Introduction B. Adrenal Cortex
I. Introduction Adrenal Glands • suprarenal – they sit on top of the kidneys • each is composed of 2 distinct regions: A. Adrenal Medulla - the inner region - comprises 20% of the gland - secretes epinephrine and norepinephrine - derived from ectoderm B. Adrenal Cortex 1) Zona Glomerulosa (outermost region) - produces mineralocorticoids (aldosterone) • the outer region 2) Zona Fasiculata (middle region) - produces glucocorticoids (cortisol) as well as • comprises 80% of the gland estrogens and androgens • secretes corticosteroids 3) Zona Reticularis (innermost region) • derived from mesoderm - same function as zona fasiculata DHEA – dehydroepiandrosterone • an adrenal androgen in females • responsible for growth of pubic and axillary hair C. Pathologies Associated with Adrenal II. Mineralocorticoids (Aldosterone) Androgen Hypersecretion A. Functions 1.Adrenogenital Syndrome - promotes reabsorption of Na+ and - hypersecretion of androgens or estrogens secretion of K+ from the distal portion of the a) in the adult female: nephron..primary regulator of salt balance and extracellular volume - masculinization (i.e. hirsutism) -Similar (but less important) effect on salt b) in the female embryo: transport in colon, salivary glands, and - female pseudohermaphroditism sweat glands. c) in the adult male: - no effect d) in young boys: - precocious pseudopuberty 1 II. Mineralocorticoids (Aldosterone) C. Pathologies B. Regulation of Secretion 1. Hypersecretion 1. Renin Angiotensin a. primary hyperaldosteronism - Angiotensin II stimulates aldost. secretion - Conn’s syndrome 2. Potassium - usually due to a tumor on the gland + - high levels of K induce aldost. secretion - too much secretion of gland itself 3. ACTH –no direct role b. secondary hyperaldosteronism - default in renin angiotensin system - most common in atherosclerosis of renal arteries C. Pathologies III. Glucocorticoids (Cortisol) 1. -
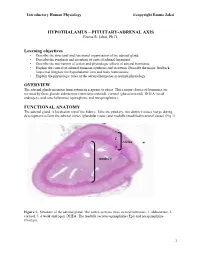
HYPOTHALAMUS – PITUITARY-ADRENAL AXIS Learning Objectives OVERVIEW FUNCTIONAL ANATOMY
Introductory Human Physiology ©copyright Emma Jakoi HYPOTHALAMUS – PITUITARY-ADRENAL AXIS Emma R. Jakoi, Ph.D. Learning objectives • Describe the structural and functional organization of the adrenal gland. • Describe the synthesis and secretion of cortical adrenal hormones. • Describe the mechanism of action and physiologic effects of adrenal hormones. • Explain the control of adrenal hormone synthesis and secretion. Describe the major feedback loops that integrate the hypothalamic axis and body homeostasis. • Explain the physiologic roles of the adrenal hormones in normal physiology. OVERVIEW The adrenal glands maintain homeostasis in response to stress. Three major classes of hormones are secreted by these glands: aldosterone (mineralocorticoid), cortisol (glucocorticoid), DHEA (weak androgen), and catecholamines (epinephrine and norepinephrine). FUNCTIONAL ANATOMY The adrenal gland is located on top of the kidney. Like the pituitary, two distinct tissues merge during development to form the adrenal cortex (glandular tissue) and medulla (modified neuronal tissue) (Fig 1). 1 2 cortex 3 medulla Figure 1. Structure of the adrenal gland. The cortex secretes three steroid hormones: 1. aldosterone, 2. cortisol, 3. a weak androgen, DHEA. The medulla secretes epinephrine (Epi) and norepinephrine (NorEpi). 1 Introductory Human Physiology ©copyright Emma Jakoi MINERALOCORTICOIDS The major mineralocorticoid in humans is aldosterone. Aldosterone is NOT under the hypothalamus- pituitary control and does not mediate a negative feedback to this axis. Aldosterone secretion is increased by the vasoconstrictor, angiotensin II, and by elevated plasma K+ concentration. Elevated plasma Na+ inhibits the secretion of aldosterone. Aldosterone, acts in the kidney to promote secretion of K+ into the urine from the blood and the reabsorption of Na+ from the urine into the blood. -

Dream Recall/Affect and the Hypothalamic–Pituitary–Adrenal Axis
Review Dream Recall/Affect and the Hypothalamic–Pituitary–Adrenal Axis Athanasios Tselebis 1, Emmanouil Zoumakis 2 and Ioannis Ilias 3,* 1 Department of Psychiatry, Sotiria Hospital, 115 27 Athens, Greece; [email protected] 2 First Department of Pediatrics, National and Kapodistrian University of Athens Medical School, Aghia Sophia Children’s Hospital, 115 27 Athens, Greece; [email protected] 3 Department of Endocrinology, Diabetes and Metabolism, Elena Venizelou Hospital, 115 21 Athens, Greece * Correspondence: [email protected]; Tel.: +30-213-205-1389 Abstract: In this concise review, we present an overview of research on dream recall/affect and of the hypothalamic–pituitary–adrenal (HPA) axis, discussing caveats regarding the action of hormones of the HPA axis (mainly cortisol and its free form, cortisol-binding globulin and glucocorticoid receptors). We present results of studies regarding dream recall/affect and the HPA axis under physiological (such as waking) or pathological conditions (such as in Cushing’s syndrome or stressful situations). Finally, we try to integrate the effect of the current COVID-19 situation with dream recall/affect vis-à-vis the HPA axis. Keywords: dreams; cortisol; stress; memory; sleep; HPA axis 1. Introduction—Dream Recall Almost all humans dream (indeed, there may be 0.5% of people who do not do so) [1]. Dreams are a series of images, thoughts and senses and are considered a specific type Citation: Tselebis, A.; Zoumakis, E.; of experience that occurs in our brain during sleep. During the dream, there is limited Ilias, I. Dream Recall/Affect and the control of the dream content, visual images and memory activation. -

Pig Gonads, Adrenal Glands and Brain C
Immunoreactive cytochrome P-45017\g=a\in rat and guinea- pig gonads, adrenal glands and brain C. Le Goascogne1, N. Sanan\l=e'\s1, M. Gou\l=e'\zou1, S. Takemori2, S. Kominami2, E. E. Baulieu1 and P. Robel1 1INSERM U33, Communications Hormonales, and Faculté de Médecine, Université Paris-sud, Lab Hormones F-94275 Bicêtre Cedex France 2 Faculty of Integrated Arts and Sciences, Hiroshima University, Hiroshima 730, Japan Summary. The cytochrome P-45017\g=a\-hydroxylase, 17\ar=r\20lyase (P-45017\g=a\) is the key enzyme responsible for the biosynthesis of androgens in steroidogenic organs. Its cellular localization has been examined with an immunohistochemical technique. In immature rat ovary, P-45017\g=a\was first detected in sparse interstitial cells on postnatal Day 8. The number of immunoreactive interstitial cells increased thereafter and the intensity of P-45017\g=a\staining in these cells was highest at 3 weeks of age. The intensity of staining then started to decline and was very faint at Day 35. From 6 weeks on, the distribution of immunoreactive P-45017\g=a\was of the adult type: it was detected exclusively in the thecal cells of the large antral, preovulatory, follicles. P-45017\g=a\was not detectable during pregnancy except on the day of parturition, when thecal cells were transiently immunoreactive. The staining had vanished 24 h after delivery. Human chorionic gonadotrophin (hCG), injected into immature females on Days 24 to 26, induced P-45017\g=a\prematurely in thecal cells. When injected on Days 12 to 14 of pregnancy, hCG also induced P-45017\g=a\in the thecal cells surrounding the largest follicles, whereas the interstitial and luteal cells were not immunostained. -

Adrenal Gland Hormones
CHAPTER 8 Adrenal Gland Hormones Devra K. Dang, PharmD, BCPS, CDE, FNAP | Trinh Pham, PharmD, BCOP | Jennifer J. Lee, PharmD, BCPS, CDE LEARNING OBJECTIVES KEY TERMS AND DEFINITIONS After completing this chapter, you should be able to ACTH (adrenocorticotropic hormone) — a hormone produced 1. Identify the hormones produced by the adrenal glands by the pituitary gland that stimulates 2. Describe the functions of mineralocorticoids and glucocorticoids in the body the adrenal cortex to produce glucocorticoids, mineralocorticoids, 3. Recognize the signs and symptoms of adrenal insuffi ciency and androgens. PART 4. Describe the pharmacological treatment of patients with acute and chronic adrenal Addison ’ s disease — a disorder insuffi ciency in which the adrenal glands do not produce enough steroid hormones. 3 5. Recognize the signs and symptoms of Cushing ’ s syndrome and the result of too Adenoma — a benign much cortisol (noncancerous) tumor of glandular 6. Describe the pharmacologic and nonpharmacologic management of patients with origin. Cushing ’ s syndrome Adrenal insuffi ciency — a term 7. List management strategies for administration of glucocorticoid and mineralocorti- referring to a defi ciency in the levels of adrenal hormones. coid therapy to avoid development of adrenal disorders Aldosterone — the hormone produced by the adrenal glands that regulates the balance of sodium, he adrenal glands are an integral part of the endocrine system, secreting water, and potassium concentrations in the body. T hormones that act throughout the body to regulate functions and promote Corticotropin-releasing homeostasis. In addition to the neurotransmitters epinephrine and norepineph- hormone (CRH) — a hormone rine, the corticosteroids secreted by the adrenal glands are vital to a wide released by the hypothalamus that variety of physiological processes. -

Cortisol-Related Signatures of Stress in the Fish Microbiome
fmicb-11-01621 July 11, 2020 Time: 15:28 # 1 ORIGINAL RESEARCH published: 14 July 2020 doi: 10.3389/fmicb.2020.01621 Cortisol-Related Signatures of Stress in the Fish Microbiome Tamsyn M. Uren Webster*, Deiene Rodriguez-Barreto, Sofia Consuegra and Carlos Garcia de Leaniz Centre for Sustainable Aquatic Research, College of Science, Swansea University, Swansea, United Kingdom Exposure to environmental stressors can compromise fish health and fitness. Little is known about how stress-induced microbiome disruption may contribute to these adverse health effects, including how cortisol influences fish microbial communities. We exposed juvenile Atlantic salmon to a mild confinement stressor for two weeks. We then measured cortisol in the plasma, skin-mucus, and feces, and characterized the skin and fecal microbiome. Fecal and skin cortisol concentrations increased in fish exposed to confinement stress, and were positively correlated with plasma cortisol. Elevated fecal cortisol was associated with pronounced changes in the diversity and Edited by: Malka Halpern, structure of the fecal microbiome. In particular, we identified a marked decline in the University of Haifa, Israel lactic acid bacteria Carnobacterium sp. and an increase in the abundance of operational Reviewed by: taxonomic units within the classes Clostridia and Gammaproteobacteria. In contrast, Heather Rose Jordan, cortisol concentrations in skin-mucus were lower than in the feces, and were not Mississippi State University, United States related to any detectable changes in the skin microbiome. Our results demonstrate that Timothy John Snelling, stressor-induced cortisol production is associated with disruption of the gut microbiome, Harper Adams University, United Kingdom which may, in turn, contribute to the adverse effects of stress on fish health. -

Posttraumatic Stress Disorder
TRAUMA AND STRESSOR RELATED DISORDERS POSTTRAUMATIC STRESS DISORDER What it is: In posttraumatic stress disorder, or PTSD, specific mental and emotional symptoms develop after an individual has been exposed to one or more traumatic events. The traumatic event experienced can range from war, as a combatant or a civilian, physical attack or assault, sexual violence, childhood physical or sexual abuse, natural disasters or a severe car accident. The traumatic events do not have to be experienced first-hand for the individual to develop PTSD, it can also develop as a result of witnessing a traumatic event, or through indirect exposure – when a traumatic event happens to a close friend or relative. Symptoms of PTSD can include distressing memories or dreams of the traumatic event, an avoidance of anything that is a reminder of the event, flashbacks of the event, as well as mood changes such as becoming more irritable, aggressive or hyper vigilant. In young children, developmental regression such as loss of language may occur. These symptoms can cause major disruptions and impairment to the individual’s ability to function at home, school and work. Individuals with PTSD are also 80% more likely to have symptoms of at least one other mental disorder, such as depressive, bipolar or substance use disorders. Common Symptoms: The following symptoms must be associated with one or more traumatic events the individual has experienced, witnessed or been indirectly exposed to. 1. Recurring and distressing memories of the event 2. Recurring and distressing dreams relating to the event 3. Dissociative reactions, such as flashbacks, in which the individual may feel or act as if the traumatic event were taking place again 4. -

Adrenal and Paraganglia Tumors Adrenal Cortical Tumors IHC: (+) SF1, Inhibin, Melan-A, Calretinin, Synaptophysin, (-) Chromogranin, Cytokeratin, S100
Last updated: 11/11/2020 Prepared by Kurt Schaberg Adrenal and Paraganglia Tumors Adrenal Cortical Tumors IHC: (+) SF1, Inhibin, Melan-A, Calretinin, Synaptophysin, (-) Chromogranin, Cytokeratin, S100. Often variable!! Adrenal Cortical Adenoma Benign. Very common. Often incidentally identified. Usually unilateral solitary masses with atrophic background adrenal gland. Tumor cells can be lipid-rich (clearer) or lipid-poor (pinker) arranged in nests and cords separated by abundant vasculature. Occasional lipofuscin pigment. Nuclei generally small and round (occasional extreme “endocrine atypia” is common). Low/no mitotic activity. Intact reticulin framework. On a spectrum with and may hard to differentiate from hyperplastic nodules, which is more often multinodular (background hyperplasia) and bilateral. Can be non-functional (85%) or functional (15%). Associated with MEN1, FAP, Carney Complex, among Aldosterone-producing→ “Conn syndrome”→ others… hypertension and hypokalemia If aldosterone-secreting adenoma is treated with Cortisol-producing→ (ACTH-independent) spironolactone→ “spironolactone bodies” (below) “Cushing Syndrome” → central obesity, moon face, hirsutism, poor healing, striae Sex-hormone-producing→ Rare (more common in carcinomas). Symptoms depend on hormone/sex (virilization or feminization) Adrenal Cortical Carcinoma Malignant. Most common in older adults. Can present with an incidental unilateral mass or with an endocrinopathy (see above). Solid, broad trabeculae, or large nested growth (more diffuse, and larger groups than in adenomas) Thick fibrous capsule with occasional fibrous bands. Frequent tumor necrosis. Frequent vascular or capsular invasion. Increased mitotic activity. Variants: Oncocytic, Myxoid, Sarcomatoid Mostly sporadic, but can be associated with Lynch Syndrome and Li-Fraumeni Syndrome Distinguishing between an Adrenal Cortical Adenoma vs Carcinoma Weiss Criteria: Weiss Criteria (≥3 = Malignant) Most widely used system, but doesn’t work as well High nuclear grade (based of Fuhrman criteria) in borderline cases or variants. -

Disturbances of the Stress Response
Disturbances of the Stress Response The Role of the HPA Axis During Alcohol Withdrawal and Abstinence Bryon Adinoff, M.D., Ali Iranmanesh, M.D., Johannes Veldhuis, M.D., and Lisa Fisher, Ph.D. Interactions among the brain, the pituitary gland, and the adrenal glands (i.e., the hypothalamic-pituitary-adrenal [HPA] axis) help regulate the body’s response to stress. The adrenal hormone cortisol plays a key role in stress reduction through its effects on multiple body systems. Excessive cortisol activity during both chronic alcohol administration and withdrawal may underlie some of the clinical complications of alcoholism, including increased risk of infectious diseases; bone, muscle, and reproductive system changes; altered energy metabolism; and disorders of mood and intellect. Despite excessive cortisol levels during intoxication and withdrawal, however, the HPA axis becomes less responsive to stress during abstinence, potentially resulting in an impaired capacity to cope with relapse-inducing stressors. KEY WORDS: AOD withdrawal syndrome; physiological stress; hypothalamic-pituitary axis; pituitary-adrenal axis; cortisol; AOD abstinence; chronic AODE (alcohol and other drug effects); corticotropin RH; arginine; vasopressin; adrenocorticotropic hormone; secretion; metabolic disorder; AODR (alcohol and other drug related) disorder; mood and affect disturbance; personality disorder; infection; drug therapy; literature review tress is a ubiquitous and unavoid- a wide range of critical physiological secretion occurs during both chronic able experience of daily life whether processes, the activity of cortisol must alcohol consumption and alcohol Sit arises from the external environ- be tightly controlled by the body. withdrawal. This heightened secretion ment (e.g., a job interview or traffic Cortisol secretion is regulated by rate may alter energy metabolism, accident) or from within the body (e.g., interactions among three structures: mental status, the structural integrity an infection or a panic attack). -

Adrenal Cortical Tumors, Pheochromocytomas and Paragangliomas
Modern Pathology (2011) 24, S58–S65 S58 & 2011 USCAP, Inc. All rights reserved 0893-3952/11 $32.00 Adrenal cortical tumors, pheochromocytomas and paragangliomas Ricardo V Lloyd Department of Pathology, University of Wisconsin School of Medicine and Public Health, Madison, WI, USA Distinguishing adrenal cortical adenomas from carcinomas may be a difficult diagnostic problem. The criteria of Weiss are very useful because of their reliance on histologic features. From a practical perspective, the most useful criteria to separate adenomas from carcinomas include tumor size, presence of necrosis and mitotic activity including atypical mitoses. Adrenal cortical neoplasms in pediatric patients are more difficult to diagnose and to separate adenomas from carcinomas. The diagnosis of pediatric adrenal cortical carcinoma requires a higher tumor weight, larger tumor size and more mitoses compared with carcinomas in adults. Pheochromocytomas are chromaffin-derived tumors that develop in the adrenal gland. Paragangliomas are tumors arising from paraganglia that are distributed along the parasympathetic nerves and sympathetic chain. Positive staining for chromogranin and synaptophysin is present in the chief cells, whereas the sustentacular cells are positive for S100 protein. Hereditary conditions associated with pheochromocytomas include multiple endocrine neoplasia 2A and 2B, Von Hippel–Lindau disease and neurofibromatosis I. Hereditary paraganglioma syndromes with mutations of SDHB, SDHC and SDHD are associated with paragangliomas and some pheochromocytomas. -

Endocrine System 4: Adrenal Glands
Copyright EMAP Publishing 2021 This article is not for distribution except for journal club use Clinical Practice Keywords Endocrine system/ Hormones/Adrenal glands Systems of life This article has been Endocrine system double-blind peer reviewed In this article... ● Endocrine functions of the adrenal glands ● Hormones of the adrenal glands ● The role of adrenal gland hormones in mediating essential physiological processes Endocrine system 4: adrenal glands Key points Authors Maria Andrade is honorary associate professor in biomedical science; There are two Zubeyde-Bayram Weston is senior lecturer in biomedical science; John Knight is adrenal glands: one associate professor in biomedical science; all at College of Human and Health located above each Sciences, Swansea University. kidney NT SELF- Abstract The endocrine system consists of glands and tissues that produce and ASSESSMENT Adrenal glands secrete hormones to regulate and coordinate vital bodily functions. This article, the Test your consist of two parts, fourth in an eight-part series on the endocrine system, explores the anatomy and knowledge. the cortex and the physiology of the adrenal glands, and describes how they regulate and coordinate After reading this medulla, which each vital physiological processes in the body through hormonal action. article go to produce different nursingtimes.net/ NTSAAdrenal hormones Citation Andrade M et al (2021) Endocrine system 4: adrenal glands. Nursing Times If you score 80% [online]; 117: 8, 54-58. or more, you will The adrenal cortex receive a certificate produces a diverse that you can use range of steroid as revalidation his eight-part series on the endo- colour and are positioned retroperito- evidence.