Sustainable Cider Apple Production
Total Page:16
File Type:pdf, Size:1020Kb

Load more
Recommended publications
-

Watergore Trial Orchard
NACM Short Report 5.5 Liz Copas 2005 WATERGORE TRIAL ORCHARD NOTE This trial site no longer exists and unfortunately some of the LA Disease Resistant seedling were grubbed out. Propagating material is still available of most of the other cultivars mentioned. SUMMARY Planted 1990 Main orchard planted to double rows N/S of Major, Ashton Bitter, Ellis Bitter and White Jersey at 18 x 8 on M25. This rootctock has proved rather too vigorous for the good soil on this site. In retrospect more effort was needed to control the early growth of these trees and induce cropping. The pruning trial [NACM 95/5/1] demonstrated some response to belated pruning to centre leader and bending or tying down strong lateral branches. This has served as a useful model for other orchards of these varieties. Planted 1995 Selected early harvesting seedlings from the Long Ashton [LA 1978] breeding program; 2 bittersweet, 4 sharps and 1 sweet, planted E/W on MM 106. Poor tree shape and excessively early flowering has ruled out many of these. The best are LA 13/2 and LA 13/7, Tremletts crosses with a strong resemblance to the parent but with some resistance to scab and mildew. Both need some initial tree training but could be kept annual. Planted 1996 Selected old varieties with some potential for bush orchards were planted on MM 106 . Of these the most promising are Broxwood Foxwhelp and possibly the other Foxwhelps [all bittersharps], also Don's Seedling [bittersweet] and Crimson King [sharp] as early harvesting varieties. Both Severn Banks [sharp] and Black Dabinett [bittersweet] could make useful late harvesting varieties. -

Paul Vossen Specialty Crops Advisor – UCCE (Sonoma-Marin)
Paul Vossen Specialty Crops Advisor – UCCE (Sonoma-Marin) http://cesonoma.ucanr.edu [email protected] Orchard Location • No Soil Problems • Full Sun • Air Drainage • Well Drained Soil • Irrigation Water • Cool Winter • 400-1,000 hrs chilling • No frost or rain during bloom (April) Soil Testing • Multiple sub samples (5-15) • Two depths (6” and 18”) (separate) • Each sample represents different area • Mix thoroughly • Analyze for big 6: pH, P, K, Ca, Mg, OM • Analyze for minor nutrients if suspect (boron, sodium, chloride, etc.) Soil Chemical Properties saturated paste extract • Soil pH 5.0 – 8.5 • High Magnesium (< 1:1 ratio with Ca) • High Calcium (> 8:1 ratio with Mg) • Adequate Phosphorous (> 10 ppm P) = OK • Adequate Potassium (> 125 ppm K) = OK • High Chloride (> 10-15 meq/l Cl-) • High Boron (> 2 ppm B) • High Sodium (SAR > 15) Soil Physical Properties Prefer OK • Sandy loam • Sandy soils • Loam • Silt loam Avoid • Clay loam • Clay soils • Silty clay loam (slow drainage) Wet soil problem one solution is Mound Planting Rootstock – Size Control M-9 2nd Year Apple Rootstocks Apple Rootstocks Apple Rootstocks Rootstock Effects ‘Yarlington Mill’ on M106 (L) and M9 (R) Cider Varieties Hard Cider Varieties (bittersweet – astringent – tannic – acidic) • Chisel Jersey • Trancendant Crab • Ashton Brown • Foxwhelp • Hyslop Crab • Kingston Black • Roxbury Russet • Pearmain • Dufflin • Yarlington Mill Cider Apple Varieties and Juice Quality C. Miles and J. King Northwestern Washington Research and Extension Center http://maritimefruit.wsu.edu -

LA GACETA N° 160 De La Fecha 24 08 2017
La Uruca, San José, Costa Rica, jueves 24 de agosto del 2017 AÑO CXXXIX Nº 160 60 páginas Estimado usuario: Si necesita consultar alguna Gaceta o Boletín Judicial anterior al año 2005 comuníquese con nuestro Archivo Institucional. 2296-9570 ext. 134 [email protected] www.imprentanacional.go.cr Pág 2 La Gaceta Nº 160 — Jueves 24 de agosto del 2017 Estado dentro del término de cinco días hábiles, contados a partir CONTENIDO de la tercera publicación de este edicto, en el Diario Ofcial La Gaceta.—San José, a las 13:20 horas del 27 de julio del 2017.— Pág Unidad de Registro de Agroquímicos y Equipos de Aplicación.— Nº Ing. Esaú Miranda Vargas, Jefe.—( IN2017159588 ). DOCUMENTOS VARIOS................................................. 2 SERVICIO NACIONAL DE SALUD ANIMAL TRIBUNAL SUPREMO DE ELECCIONES DIRECCION GENERAL Edictos ............................................................................. 40 DIRECTRIZ SENASA-DG-D001-2017 Avisos .............................................................................. 40 Considerando: CONTRATACIÓN ADMINISTRATIVA ....................... 41 I.—Que de conformidad con el inciso e) del artículo 6 de la Ley REGLAMENTOS ............................................................ 42 N° 8495 del 6 de abril del 2006, Ley General del Servicio Nacional de Salud Animal (SENASA), es competencia de dicho Servicio INSTITUCIONES DESCENTRALIZADAS ................ 44 “Dictar las normas técnicas pertinentes, elaborar los manuales RÉGIMEN MUNICIPAL ................................................ 47 de -

National Apple Orchard Census 2012
National Apple Orchard Census 2012 1 Table of Contents Introduction Page 3 Executive Summary Page 4 Section 1 Orchard Area, Sales, Growers and Markets Page 6 Section 2 Varieties and Orchard Age Page 15 Section 3 Employment, Facilities, Business Development and Producer Page 21 Profile Appendix Tables Table 1: Apple Production Area (hectares) by Year of Planting and Apple Type in Ireland in 2012 Table 2: Number of Apple Trees by Year of Planting and Apple Type in Ireland in 2012 Table 3: Number of Growers of Each Apple Type by County in Ireland in 2012 Table 4: Sales of 2011 Harvest by Market by County Table 5: Production Area, Number of Growers and Sales for Most Commonly Grown Apple Varieties in Ireland in 2011/12 Table 6: Average (median) Prices per Tonne by Market for 2011 harvest Table 7: Apple Production Area (hectares) in Ireland in 1980-2012 2 National Apple Orchard Census 2012 Introduction The National Apple Orchard Census 2012 was compiled from information provided by the 45 commercial apple growers operating in the Republic of Ireland in 2012. The census provides an important overview of apple production, and comparisons are made to previous censuses (2007, 2002 and 1997) where possible. The census surveyed all commercial orchards, defined as follows: - a production area of at least 0.2 hectares of apples, including cider apples - the fruit from the orchard is sold every year - the orchard is sprayed against pests/diseases every year, or maintained to organic standards Within the 45 growers, there are two organic growers. Production areas are based on areas planted up to March 2012. -

A Big Win for the Msd: Mackey Pendergrast Named Superintendent of the Year for the State of New Jersey!
A joint publication from The Morris School District and The Morris Educational Foundation FALL 2019 FIND OUT WHAT’S HAPPENING IN... DTHE istrict A BIG WIN FOR THE MSD: MACKEY PENDERGRAST NAMED SUPERINTENDENT OF THE YEAR FOR THE STATE OF NEW JERSEY! The New Jersey Association of School Administrators (NJASA) has chosen Mackey Pendergrast, Superintendent of the Morris School District, for New Jersey’s Superintendent of the Year. Mr. Pendergrast will now represent the State as a candidate for the American Association of School Administrators (AASA) National Superintendent of the Year Award, which recognizes the talent, vision, and impact of our nation’s public school leaders. He was named Morris County Superintendent of the Year in June and Region I Superintendent of the Year for North Jersey in September. This distinction comes at the start of Pendergrast’s fifth year as superintendent of the Morris School District and his ninth year overall as a superintendent. He began his public school teaching career as a high school history teacher and varsity basketball coach for the Summit Public Schools and held positions in central office administration for the Westfield Public School District and the West Morris Regional High School District before becoming superintendent of West Morris in 2011. Board of Education President Lisa Pollak and Vice President Nancy Bangiola said they were “absolutely thrilled but hardly surprised” by their superintendent’s recent accolades, including the state’s top honors. more on page 3> MEF and MSD Family Author Experience Kicks Off Love of Reading What do writers and ninjas have in common? District second graders contemplated this question in October during the fall installment of the new Morris Educational Foundation (MEF) and Morris School District (MSD) Family Author Experience. -

Survey of Apple Clones in the United States
Historic, archived document Do not assume content reflects current scientific knowledge, policies, or practices. 5 ARS 34-37-1 May 1963 A Survey of Apple Clones in the United States u. S. DFPT. OF AGRffini r U>2 4 L964 Agricultural Research Service U.S. DEPARTMENT OF AGRICULTURE PREFACE This publication reports on surveys of the deciduous fruit and nut clones being maintained at the Federal and State experiment stations in the United States. It will b- published in three c parts: I. Apples, II. Stone Fruit. , UI, Pears, Nuts, and Other Fruits. This survey was conducted at the request of the National Coor- dinating Committee on New Crops. Its purpose is to obtain an indication of the volume of material that would be involved in establishing clonal germ plasm repositories for the use of fruit breeders throughout the country. ACKNOWLEDGMENT Gratitude is expressed for the assistance of H. F. Winters of the New Crops Research Branch, Crops Research Division, Agricultural Research Service, under whose direction the questionnaire was designed and initial distribution made. The author also acknowledges the work of D. D. Dolan, W. R. Langford, W. H. Skrdla, and L. A. Mullen, coordinators of the New Crops Regional Cooperative Program, through whom the data used in this survey were obtained from the State experiment stations. Finally, it is recognized that much extracurricular work was expended by the various experiment stations in completing the questionnaires. : CONTENTS Introduction 1 Germany 298 Key to reporting stations. „ . 4 Soviet Union . 302 Abbreviations used in descriptions .... 6 Sweden . 303 Sports United States selections 304 Baldwin. -

We Make Delicious Cider from Freshly-Crushed Fruit How We Make It Happen the Zeffer Story
WE MAKE DELICIOUS CIDER FROM FRESHLY-CRUSHED FRUIT HOW WE MAKE IT HAPPEN THE ZEFFER STORY OUR JOURNEY BEGAN ON SAM’S PARENTS FARM IN 2009 WHEN SAM DECIDED TO TRY HIS WINE-MAKING HAND AT MAKING CIDER. AFTER EXTENSIVE RESEARCH WE KNEW THE STYLE OF CIDER WE LIKED BEST. NOT JUST ANY OLD ‘MADE FROM CONCENTRATE’ CIDER, WE WANTED TO MAKE REAL CIDER FROM REAL FRUIT WITH PATIENCE, CRAFT AND QUALITY. We knew that the final product would taste its best if we started with the best ingredients so we scoured the country to find specific apple and pear varieties from orchards around New Zealand. After long wintery nights crushing, an exploding fruit press, experimental brews and many hours spent hand bottling we had our first batch ready for release in the Spring of 2009. We sold it exclusively through our local Matakana Farmers market and were rewarded with great feedback, eager buyers and steady growth which ultimately allowed us to build our own cidery. And, while we are now the proud owners of a shiny new fruit press we remain faithful to our simple vision and ethos of making cider that we are both proud to put our name on and love to drink. Our ciders are fermented in small batches and made with minimal intervention that allows the natural flavours and true character to shine without the use of any artificial colours or sweeteners. We love what we do and we love that we get to share it with you. From the Team at Zeffer PRODUCT RANGE We make a tasty drop or two Or for something a little more fancy Red Apple Cider FRESH FROM THE ORCHARD Our Red Apple Cider captures the fresh flavour of the unique Mahana Red apples used to make it. -

January 22, 2020 Director General, Telecommunications and Internet
Denis Marquis President, Bell Pensioners' Group 1914, rue Cugnet Saint-Bruno de Montarville QC J3V 5H6 Tel.: 450 441-0111 Email: [email protected] January 22, 2020 Director General, Telecommunications and Internet Policy Branch, Innovation, Science and Economic Development Canada, 235 Queen Street, 10th Floor, Ottawa, Ontario K1A 0H5 Sent via e-mail to: [email protected] Subject: Canada Gazette, Part I, December 14, 2019, Volume 153, Number 10, Notice No. TIPB-002-2019 — Petitions to the Governor in Council concerning Telecom Order CRTC 2019-288 Dear Madam: Attached please find a submission by the Bell Pensioners’ Group (BPG) concerning the above- noted matter. Yours truly, Denis Marquis President, Bell Pensioners’ Group www.bellpensionersgroup.ca Attachment c.c: Hon. Navdeep Bains, P.C., M.P., Minister of Innovation, Science and Industry [email protected] Hon. Deb Schulte, P.C., M.P., Minister of Seniors [email protected] Attachment Canada Gazette, Part I, December 14, 2019, Volume 153, Number 10, Notice No. TIPB-002-2019 — Petitions to the Governor in Council concerning Telecom Order CRTC 2019-288 Comments of the Bell Pensioners’ Group In accordance with Gazette Notice No. TIPB-002-2019, the Bell Pensioners’ Group (BPG) is pleased to provide this submission in support of a petition to the Governor in Council by Bell Canada seeking a variation of a decision issued by the Canadian Radio-television and Tele- communications Commission (CRTC) concerning final rates for aggregated wholesale high- speed internet access services. This CRTC decision requires that facilities-based competitors like Bell Canada (Bell) allow use of their state-of-the-art communications networks by resale- based competitors at substantially reduced prices that are, in the case of Bell, below its costs incurred to build those networks. -

The Craft Cider Revival – Some Technical Considerations Andrew Lea 28/2/2007 1
The Craft Cider Revival – Some Technical Considerations Andrew Lea 28/2/2007 1 THE CRAFT CIDER REVIVAL ~ Some Technical Considerations Presentation to SWECA 28th February 2007 Andrew Lea SOME THINGS TO THINK ABOUT Orcharding and fruit selection Full juice or high gravity fermentations Yeast and sulphiting Keeving Malo-lactic maturation Style of finished product What is your overall USP? How are you differentiated? CRAFT CIDER IS NOW SPREADING Cidermaking was once widespread over the whole of Southern England There are signs that it may be returning eg Kent, Sussex and East Anglia So regional styles may be back in favour eg higher acid /less tannic in the East CHOICE OF CIDER FRUIT The traditional classification (Barker, LARS, 1905) Acid % ‘Tannin’ % Sweet < 0.45 < 0.2 Sharp > 0.45 < 0.2 Bittersharp > 0.45 > 0.2 Bittersweet < 0.45 > 0.2 Finished ~ 0.45 ~ 0.2 Cider CHOICE OF “VINTAGE QUALITY” FRUIT Term devised by Hogg 1886 Adopted by Barker 1910 to embrace superior qualities that could not be determined by analysis This is still true today! The Craft Cider Revival – Some Technical Considerations Andrew Lea 28/2/2007 2 “VINTAGE QUALITY” LIST (1988) Sharps / Bittersharps Dymock Red Kingston Black Stoke Red Foxwhelp Browns Apple Frederick Backwell Red Bittersweets Ashton Brown Jersey Harry Masters Jersey Dabinett Major White Jersey Yarlington Mill Medaille d’Or Pure Sweets Northwood Sweet Alford Sweet Coppin BLENDING OR SINGLE VARIETALS? Blending before fermentation can ensure good pH control (< 3.8) High pH (bittersweet) juices prone to infection Single varietals may be sensorially unbalanced unless ameliorated with dilution or added acid RELATIONSHIP BETWEEN pH AND TITRATABLE ACID IS NOT EXACT Most bittersweet juices are > pH 3.8 or < 0.4% titratable acidity. -

Brown Snout’ Specialty Cider Apple U.S
most popular alcoholic beverage made Yield, Labor, and Fruit and Juice Quality andconsumedintheUnitedStates; Characteristics of Machine and Hand-harvested however, by the early 1900s, cider had essentially disappeared from ‘Brown Snout’ Specialty Cider Apple U.S. markets (Proulx and Nichols, 1997). The rapid decline of cider 1 was due to a combination of factors, Carol A. Miles and Jaqueline King primarily a high influx of German and eastern European immigrants who ADDITIONAL INDEX WORDS. fruit storage, hard cider, harvest labor, Malus ·domestica, preferred beer, and many farmers mechanical fruit harvest, over-the-row harvester who were sympathetic to the Tem- perance Movement cut down their SUMMARY. In this 2-year study of ‘Brown Snout’ specialty cider apple apple trees (Watson, 1999). (Malus ·domestica) grafted onto Malling 27 (M.27) and East Malling/Long Ashton Cider is currently seeing a revival 9, we compared weight of total harvested fruit, labor hours for harvest, tree and fruit damage, and fruit and juice quality characteristics for machine and hand harvest. in the United States and although it Machine harvest was with an over-the-row small fruit harvester. There were no only accounts for 1% of the alcoholic significant differences due to rootstock; however, there were differences between beverage market, it is the fastest years for most measurements. Weight of harvested fruit did not differ because of growing alcohol market segment, harvest method; however, harvest efficiency was 68% to 72% for machine pick and with 54% increase in production each 85% to 89% for machine pick D clean-up weight (fruit left on trees and fruit year from 2007 to 2012 (Morton, knocked to the ground during harvest) as compared with hand harvest. -
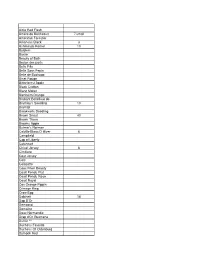
Scion List 2013 April 6
Airlie Red Flesh Amere de Berthcourt 7 small American Forestier Arkansas Black 3 Ashmeads Kernel 10 Baldwin Baxter Beauty of Bath Bedan des parts Belle Fille Belle Sans Pepin Belle de Boskoop Binet Rouge Bitterforest Apple Black Crofton Blanc Mollet Blenheim Orange Brabant Bellefleur de Bramley's Seedling 10 Bramtot Breakwells Seedling Brown Snout 40 Brown Thorn Browns Apple Bulmer's Norman Calville Blanc D Hiver 6 Campfield Cap of Liberty Catshead Chisel Jersey 6 Cimitiere Coat Jersey Cole Colozette Coos River Beauty Court Pendu Plat Court Pendu Rose Court Royal Cox Orange Pippin Crimson King Crow Egg Dabinett 18 Dap D'Or Democrat Domaine Doux Normandie Drap d'Or Guemene Dufflin ** Duchess Favorite Duchess Of Oldenburg Dymock Red Ellis Bitter English Redstreak Esopus Spitzenburg Fall Pippin Fameuse Fillbarrel Finkenwerder Herbstprinz Flower of Kent Foxwhelp Frequin Frequin Audievre Geeveston Fanny 6 Gilpin Golden Harvey Golden Hornet Golden Noble Golden Russett 35 Goolsbey Granniwinkle Granny Smith Grimes Golden Grindstone Gros Frequin Harrison Harrys Master Jersey 7 Harvey Holland Improved Dove Karmijn De Sonneville Keswick Codlin Kidd's Orange Red 4 Kingston Black Lambrook Pippin Le Bret Liberty London Pippin Lord Derby Loyalist Lubsk Queen Lyman Prolific ** Major Marachal Marin Onfroy Maude Medaille D'Or Melrose Merton Russet Metais Mettais Michelin 13 Minnesota 1734 Muscadet de Dieppe 11 Muscadet de Lense Muscadet des Bernet Nehou Noel Deschamps Norfolk Beefing Northern Spy Northwood Peau D'Ane Peau de Vache Pethyre Pitmaston Pine Apple Plum Pomme Gris ** Porters Perfection Red Astrachan Red Cox's Pamona Red Jersey (loyal drain) Red Spitzenburg Redstreak Reine des Hatives Reine des Pommes 11 Reinette de Reinette 3 Reinette Du Canada Reinette Gris 1 Rene Martin Ribston Pippin Rhode Island Greenling Ross Nonpareil Rousse Latour Roxbury Russett Sherrington Norman ** Skyrmes Kernel Slippery Cider Somerset Redstreak Souvenir de Fernand Cognet St. -

Britain's Cider Apples
eat rows of young apple trees stand in a peaceful orchard close to Hereford. THE MAN Carefully staked and protected by wire fencing, WHO SAVED they guide the eye towards spectacular views of Hay Bluff and the Black Mountains. This is Tidnor Wood, Henry NMay’s piece of paradise, where for 14 years he worked, worried and enjoyed many moments BRITAIN’S of pleasure – picking apples as they hung heavy on the boughs, sharing barbecues with workers at the end of a hard day. Henry, however, is no ordinary grower and CIDER this is no ordinary orchard. It has now become a gene pool, regenerating Britain with hundreds of rare cider-apple varieties that would otherwise have been lost to the nation. Over years, the former Customs & Excise APPLES officer, now 71, built up a unique collection. Henry May had just been looking Slack-Ma-Girdle, Bloody Turk and Greasy Butcher are just some of the 400 varieties for a quirky project to occupy his time. growing here, with names evoking long- He ended up making an enormous vanished days when every West Country farmhouse had a few apple trees and workers contribution to our horticultural history were part-paid in cider. Yet the orchard came about almost by accident. Words Susannah Hickling In his mid-50s, Henry was living in Portrait Sean Malyon Northampton, had made money on property and wanted an unusual project. He had once been based in Hereford, where he checked the excise-duty records of cider companies and had enjoyed being involved with their work – including drinking the fruits of production! Since the Fifties, 90% of traditional orchards had vanished, lost to housing, other crops or intensive, commercial growing practices.