Cortical Representation of Tympanic Membrane Movements Due to Pressure Variation: an Fmri Study
Total Page:16
File Type:pdf, Size:1020Kb

Load more
Recommended publications
-
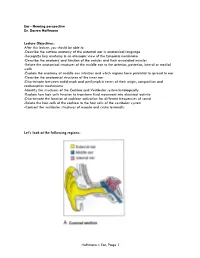
Ear, Page 1 Lecture Outline
Ear - Hearing perspective Dr. Darren Hoffmann Lecture Objectives: After this lecture, you should be able to: -Describe the surface anatomy of the external ear in anatomical language -Recognize key anatomy in an otoscopic view of the tympanic membrane -Describe the anatomy and function of the ossicles and their associated muscles -Relate the anatomical structures of the middle ear to the anterior, posterior, lateral or medial walls -Explain the anatomy of middle ear infection and which regions have potential to spread to ear -Describe the anatomical structures of the inner ear -Discriminate between endolymph and perilymph in terms of their origin, composition and reabsorption mechanisms -Identify the structures of the Cochlea and Vestibular system histologically -Explain how hair cells function to transform fluid movement into electrical activity -Discriminate the location of cochlear activation for different frequencies of sound -Relate the hair cells of the cochlea to the hair cells of the vestibular system -Contrast the vestibular structures of macula and crista terminalis Let’s look at the following regions: Hoffmann – Ear, Page 1 Lecture Outline: C1. External Ear Function: Amplification of Sound waves Parts Auricle Visible part of external ear (pinna) Helix – large outer rim Tragus – tab anterior to external auditory meatus External auditory meatus Auditory Canal/External Auditory Meatus Leads from Auricle to Tympanic membrane Starts cartilaginous, becomes bony as it enters petrous part of temporal bone Earwax (Cerumen) Complex mixture -

Bedside Neuro-Otological Examination and Interpretation of Commonly
J Neurol Neurosurg Psychiatry: first published as 10.1136/jnnp.2004.054478 on 24 November 2004. Downloaded from BEDSIDE NEURO-OTOLOGICAL EXAMINATION AND INTERPRETATION iv32 OF COMMONLY USED INVESTIGATIONS RDavies J Neurol Neurosurg Psychiatry 2004;75(Suppl IV):iv32–iv44. doi: 10.1136/jnnp.2004.054478 he assessment of the patient with a neuro-otological problem is not a complex task if approached in a logical manner. It is best addressed by taking a comprehensive history, by a Tphysical examination that is directed towards detecting abnormalities of eye movements and abnormalities of gait, and also towards identifying any associated otological or neurological problems. This examination needs to be mindful of the factors that can compromise the value of the signs elicited, and the range of investigative techniques available. The majority of patients that present with neuro-otological symptoms do not have a space occupying lesion and the over reliance on imaging techniques is likely to miss more common conditions, such as benign paroxysmal positional vertigo (BPPV), or the failure to compensate following an acute unilateral labyrinthine event. The role of the neuro-otologist is to identify the site of the lesion, gather information that may lead to an aetiological diagnosis, and from there, to formulate a management plan. c BACKGROUND Balance is maintained through the integration at the brainstem level of information from the vestibular end organs, and the visual and proprioceptive sensory modalities. This processing takes place in the vestibular nuclei, with modulating influences from higher centres including the cerebellum, the extrapyramidal system, the cerebral cortex, and the contiguous reticular formation (fig 1). -

Hearing Loss, Vertigo and Tinnitus
HEARING LOSS, VERTIGO AND TINNITUS Jonathan Lara, DO April 29, 2012 Hearing Loss Facts S Men are more likely to experience hearing loss than women. S Approximately 17 percent (36 million) of American adults report some degree of hearing loss. S About 2 to 3 out of every 1,000 children in the United States are born deaf or hard-of-hearing. S Nine out of every 10 children who are born deaf are born to parents who can hear. Hearing Loss Facts S The NIDCD estimates that approximately 15 percent (26 million) of Americans between the ages of 20 and 69 have high frequency hearing loss due to exposure to loud sounds or noise at work or in leisure activities. S Only 1 out of 5 people who could benefit from a hearing aid actually wears one. S Three out of 4 children experience ear infection (otitis media) by the time they are 3 years old. Hearing Loss Facts S There is a strong relationship between age and reported hearing loss: 18 percent of American adults 45-64 years old, 30 percent of adults 65-74 years old, and 47 percent of adults 75 years old or older have a hearing impairment. S Roughly 25 million Americans have experienced tinnitus. S Approximately 4,000 new cases of sudden deafness occur each year in the United States. Hearing Loss Facts S Approximately 615,000 individuals have been diagnosed with Ménière's disease in the United States. Another 45,500 are newly diagnosed each year. S One out of every 100,000 individuals per year develops an acoustic neurinoma (vestibular schwannoma). -

Initial Stage of Fetal Development of the Pharyngotympanic Tube Cartilage with Special Reference to Muscle Attachments to the Tube
Original Article http://dx.doi.org/10.5115/acb.2012.45.3.185 pISSN 2093-3665 eISSN 2093-3673 Initial stage of fetal development of the pharyngotympanic tube cartilage with special reference to muscle attachments to the tube Yukio Katori1, Jose Francisco Rodríguez-Vázquez2, Samuel Verdugo-López2, Gen Murakami3, Tetsuaki Kawase4,5, Toshimitsu Kobayashi5 1Division of Otorhinolaryngology, Sendai Municipal Hospital, Sendai, Japan, 2Department of Anatomy and Embryology II, Faculty of Medicine, Complutense University, Madrid, Spain, 3Division of Internal Medicine, Iwamizawa Kojin-kai Hospital, Iwamizawa, 4Laboratory of Rehabilitative Auditory Science, Tohoku University Graduate School of Biomedical Engineering, 5Department of Otolaryngology-Head and Neck Surgery, Tohoku University Graduate School of Medicine, Sendai, Japan Abstract: Fetal development of the cartilage of the pharyngotympanic tube (PTT) is characterized by its late start. We examined semiserial histological sections of 20 human fetuses at 14-18 weeks of gestation. As controls, we also observed sections of 5 large fetuses at around 30 weeks. At and around 14 weeks, the tubal cartilage first appeared in the posterior side of the pharyngeal opening of the PTT. The levator veli palatini muscle used a mucosal fold containing the initial cartilage for its downward path to the palate. Moreover, the cartilage is a limited hard attachment for the muscle. Therefore, the PTT and its cartilage seemed to play a critical role in early development of levator veli muscle. In contrast, the cartilage developed so that it extended laterally, along a fascia-like structure that connected with the tensor tympani muscle. This muscle appeared to exert mechanical stress on the initial cartilage. -

Tonic Tensor Tympani Syndrome (TTTS)
Tonic Tensor Tympani Syndrome (TTTS) http://www.dineenandwestcott.com.au/hyperacusis.php?fid=1 Retrieved 15ththth May 2009 In the middle ear, the tensor tympani muscle and the stapedial muscle contract to tighten the middle ear bones (the ossicles) as a reaction to loud, potentially damaging sound. This provides protection to the inner ear from these loud sounds. In many people with hyperacusis, an increased, involuntary activity can develop in the tensor tympani muscle in the middle ear as part of a protective and startle response to some sounds. This lowered reflex threshold for tensor tympani contraction is activated by the perception/anticipation of sudden, unexpected, loud sound, and is called tonic tensor tympani syndrome (TTTS). In some people with hyperacusis, it appears that the tensor tympani muscle can contract just by thinking about a loud sound. Following exposure to intolerable sounds, this heightened contraction of the tensor tympani muscle: • tightens the ear drum • stiffens the middle ear bones (ossicles) • can lead to irritability of the trigeminal nerve, which innervates the tensor tympani muscle; and to other nerves supplying the ear drum • can affect the airflow into the middle ear. The tensor tympani muscle functions in coordination with the tensor veli palatini muscle. When we yawn or swallow, these muscles work together to open the Eustachian tube. This keeps the ears healthy by clearing the middle ear of any accumulated fluid and allows the ears to “pop” by equalising pressure caused by altitude changes. TTTS can lead to a range of symptoms in and around the ear(s): ear pain; pain in the jaw joint and down the neck; a fluttering sensation in the ear; a sensation of fullness in the ear; burning/numbness/tingling in and around the ear; unsteadiness; distorted hearing. -

ANATOMY of EAR Basic Ear Anatomy
ANATOMY OF EAR Basic Ear Anatomy • Expected outcomes • To understand the hearing mechanism • To be able to identify the structures of the ear Development of Ear 1. Pinna develops from 1st & 2nd Branchial arch (Hillocks of His). Starts at 6 Weeks & is complete by 20 weeks. 2. E.A.M. develops from dorsal end of 1st branchial arch starting at 6-8 weeks and is complete by 28 weeks. 3. Middle Ear development —Malleus & Incus develop between 6-8 weeks from 1st & 2nd branchial arch. Branchial arches & Development of Ear Dev. contd---- • T.M at 28 weeks from all 3 germinal layers . • Foot plate of stapes develops from otic capsule b/w 6- 8 weeks. • Inner ear develops from otic capsule starting at 5 weeks & is complete by 25 weeks. • Development of external/middle/inner ear is independent of each other. Development of ear External Ear • It consists of - Pinna and External auditory meatus. Pinna • It is made up of fibro elastic cartilage covered by skin and connected to the surrounding parts by ligaments and muscles. • Various landmarks on the pinna are helix, antihelix, lobule, tragus, concha, scaphoid fossa and triangular fossa • Pinna has two surfaces i.e. medial or cranial surface and a lateral surface . • Cymba concha lies between crus helix and crus antihelix. It is an important landmark for mastoid antrum. Anatomy of external ear • Landmarks of pinna Anatomy of external ear • Bat-Ear is the most common congenital anomaly of pinna in which antihelix has not developed and excessive conchal cartilage is present. • Corrections of Pinna defects are done at 6 years of age. -

Nomina Histologica Veterinaria, First Edition
NOMINA HISTOLOGICA VETERINARIA Submitted by the International Committee on Veterinary Histological Nomenclature (ICVHN) to the World Association of Veterinary Anatomists Published on the website of the World Association of Veterinary Anatomists www.wava-amav.org 2017 CONTENTS Introduction i Principles of term construction in N.H.V. iii Cytologia – Cytology 1 Textus epithelialis – Epithelial tissue 10 Textus connectivus – Connective tissue 13 Sanguis et Lympha – Blood and Lymph 17 Textus muscularis – Muscle tissue 19 Textus nervosus – Nerve tissue 20 Splanchnologia – Viscera 23 Systema digestorium – Digestive system 24 Systema respiratorium – Respiratory system 32 Systema urinarium – Urinary system 35 Organa genitalia masculina – Male genital system 38 Organa genitalia feminina – Female genital system 42 Systema endocrinum – Endocrine system 45 Systema cardiovasculare et lymphaticum [Angiologia] – Cardiovascular and lymphatic system 47 Systema nervosum – Nervous system 52 Receptores sensorii et Organa sensuum – Sensory receptors and Sense organs 58 Integumentum – Integument 64 INTRODUCTION The preparations leading to the publication of the present first edition of the Nomina Histologica Veterinaria has a long history spanning more than 50 years. Under the auspices of the World Association of Veterinary Anatomists (W.A.V.A.), the International Committee on Veterinary Anatomical Nomenclature (I.C.V.A.N.) appointed in Giessen, 1965, a Subcommittee on Histology and Embryology which started a working relation with the Subcommittee on Histology of the former International Anatomical Nomenclature Committee. In Mexico City, 1971, this Subcommittee presented a document entitled Nomina Histologica Veterinaria: A Working Draft as a basis for the continued work of the newly-appointed Subcommittee on Histological Nomenclature. This resulted in the editing of the Nomina Histologica Veterinaria: A Working Draft II (Toulouse, 1974), followed by preparations for publication of a Nomina Histologica Veterinaria. -

Yagenich L.V., Kirillova I.I., Siritsa Ye.A. Latin and Main Principals Of
Yagenich L.V., Kirillova I.I., Siritsa Ye.A. Latin and main principals of anatomical, pharmaceutical and clinical terminology (Student's book) Simferopol, 2017 Contents No. Topics Page 1. UNIT I. Latin language history. Phonetics. Alphabet. Vowels and consonants classification. Diphthongs. Digraphs. Letter combinations. 4-13 Syllable shortness and longitude. Stress rules. 2. UNIT II. Grammatical noun categories, declension characteristics, noun 14-25 dictionary forms, determination of the noun stems, nominative and genitive cases and their significance in terms formation. I-st noun declension. 3. UNIT III. Adjectives and its grammatical categories. Classes of adjectives. Adjective entries in dictionaries. Adjectives of the I-st group. Gender 26-36 endings, stem-determining. 4. UNIT IV. Adjectives of the 2-nd group. Morphological characteristics of two- and multi-word anatomical terms. Syntax of two- and multi-word 37-49 anatomical terms. Nouns of the 2nd declension 5. UNIT V. General characteristic of the nouns of the 3rd declension. Parisyllabic and imparisyllabic nouns. Types of stems of the nouns of the 50-58 3rd declension and their peculiarities. 3rd declension nouns in combination with agreed and non-agreed attributes 6. UNIT VI. Peculiarities of 3rd declension nouns of masculine, feminine and neuter genders. Muscle names referring to their functions. Exceptions to the 59-71 gender rule of 3rd declension nouns for all three genders 7. UNIT VII. 1st, 2nd and 3rd declension nouns in combination with II class adjectives. Present Participle and its declension. Anatomical terms 72-81 consisting of nouns and participles 8. UNIT VIII. Nouns of the 4th and 5th declensions and their combination with 82-89 adjectives 9. -

Audiometric Findings with Voluntary Tensor Tympani Contraction Brandon Wickens1 , Duncan Floyd2 and Manohar Bance3*
Wickens et al. Journal of Otolaryngology - Head and Neck Surgery (2017) 46:2 DOI 10.1186/s40463-016-0182-y ORIGINALRESEARCHARTICLE Open Access Audiometric findings with voluntary tensor tympani contraction Brandon Wickens1 , Duncan Floyd2 and Manohar Bance3* Abstract Background: Tensor tympani contraction may have a "signature" audiogram. This study demonstrates audiometric findings during voluntary tensor tympani contraction. Methods: Five volunteers possessing the ability to voluntarily contract their tensor tympani muscles were identified and enrolled. Tensor tympani contraction was confirmed with characteristic tympanometry findings. Study subjects underwent conventional audiometry. Air conduction and bone conduction threshold testing was performed with and without voluntary tensor tympani contraction. Main outcome measure: Changes in air conduction and bone conduction thresholds during voluntary tensor tympani contraction. Results: Audiometric results demonstrate a low frequency mixed hearing loss resulting from tensor tympani contraction. Specifically, at 250 Hz, air conduction thresholds increased by 22 dB and bone conduction thresholds increased by 10 dB. Conclusions: Previous research has demonstrated a low frequency conductive hearing loss in the setting of tensor tympanic contraction. This is the first study to demonstrate a low frequency mixed hearing loss associated with tensor tympani contraction. This finding may aid in the diagnosis of disorders resulting from abnormal tensor tympani function. Tensor tympani contraction -

Positive Perilymph Fistula Test with Semicircular Canal Dehiscence from Cholesteatoma
PRACTICE | CLINICAL IMAGES Positive perilymph fistula test with semicircular canal dehiscence from cholesteatoma Ming-Chih Hsieh MD, Chen Chi Wu MD PhD, Shih-Hao Wang MD n Cite as: CMAJ 2019 January 28;191:E104. doi: 10.1503/cmaj.180799 54-year-old man presented to our outpatient department with left-side hearing loss and tinnitus that had progressed for several years. The patient had vertigo with nausea, whichA was aggravated on applying pressure over the left external ear canal and tragus. Physical examination showed left-side tym- panic membrane retraction with a whitish mass at the epitympa- num, suggestive of cholesteatoma. Gently compressing the left-ear tragus induced apparently left-beating horizontal nystagmus (see video, Appendix 1, available at www.cmaj.ca/lookup/suppl/ doi:10.1503/cmaj.180799/-/DC1), consistent with a positive peri- lymph fistula test. Pure tone audiometry showed mixed-type hear- ing loss of 104 dB in the patient’s left ear and sensorineural hearing loss of 62 dB in his right. High-resolution computed tomography (CT) scan of the patient’s temporal bone showed a soft-tissue mass in his left middle ear and mastoid cavity with left lateral semicircular canal erosion (Appendix 2, available at www.cmaj.ca/lookup/suppl/ Figure 1: Microscopic view of left lateral semicircular canal dehiscence with doi:10.1503/cmaj.180799/-/DC1). These findings were compatible erosion of bony and membranous sections (arrows) in a 54-year-old man with with cholesteatoma with lateral semicircular canal dehiscence. cholesteatoma. Note: *the malleus handle; +second genu of the facial nerve; During surgery, we noted that the osseous and membranous por- dotted lines define the tympanic segment of the facial nerve. -

Patient Information – Ear Surgery Instructions
The Oregon Clinic, Plaza ENT Division 5050 NE Hoyt #655, Portland, OR 97213 Phone: 5034882400 Fax: 5032310121 Patient Information – Ear Surgery Instructions Pre- and Post-operative Instructions for Ear Surgery (not including ear tubes) Before Surgery: Many ear surgeries involve manipulation of the eardrum (tympanic membrane), and some require the removal of bone to facilitate the treatment of your ear disease. As with any operation, infection, scarring, and blood clot formation (hematoma) are possible. The facial nerve is at risk for injury or temporary weakness during any ear surgery. Dizziness following surgery may be expected. Hearing loss or ringing in the ear (tinnitus) may be more pronounced. Taste disturbance is not uncommon in certain ear surgeries for a few weeks following surgery and, in a few instances, could be prolonged or permanent. An incision may be made behind your ear, on your earlobe, or behind the pointed cartilage in front of your ear (the tragus). These areas normally heal without problems or obvious scars. Hair around the ear may or may not be shaved. Flying is usually permitted one month after surgery. Swimming may be allowed six weeks after surgery, but check with your doctor first before resuming swimming or other water sports. If your work is not strenuous and depending upon the type of surgery you’ve had, you may return to work 3 to 4 days from the date of surgery. Generally, you will be seen about 2-3 weeks after surgery. This gives your eardrum time to heal before we see you back. Pre-operative Instructions: 1. -

Rare and Multiple Variation of the Auricula: (Case Report)
Global Advanced Research Journal of Medicine and Medical Sciences (ISSN: 2315-5159) Vol. 6(8) pp. 172-174, August, 2017 Available online http://garj.org/garjmms Copyright © 2017 Global Advanced Research Journals Case Report Rare and Multiple Variation of the Auricula: (Case Report) Kosif R* and Dıramalı M* Department of Anatomy, Faculty of Medicine, Abant Izzet Baysal University, Bolu, Turkey. Accepted 10 August, 2017 Ear marks and measurements are often used for identification in the absence of valid fingerprints. Auricular variations were identified in a 22-year-old male university student. Both ears were accompanied by absence of tragus, thick crus helicis, prominent antitragus, long incisura intertragica and downward-sloping convex crus antihelicis. He does not have a known disease. Auricular variations are seen 2% in normal people and 98% in genetic diseases. Keywords : Variation, Auricula, Tragus, Antitragus INTRODUCTION The human ear is divided into external, middle and structure of the external ear is utilized for personal internal parts. Auricle and external acoustic meatus form identification of living subjects in relation to criminal the external ear which is utilized in the forensic sciences activity (Swift and Rutty, 2003 ). for individual identification and authentication (Purkait Normal auricular anatomy has been showed in Figure 1. and Singh, 2007). The lateral surface of the auricle is irregularly concave, faces slightly forward and displays many eminences and depressions, which can make contact various surfaces and can produce a print as a rubber stamp (Meijerman et al., 2004). Ear marks and measurements are often used for identification in the absence of valid fingerprints. The dimensions of the pinna have been found to vary among different ethnic groups (Kumar and Singla, 2013).