Global Correlation Analysis for Micro-RNA and Mrna Expression Profiles in Human Cell Lines
Total Page:16
File Type:pdf, Size:1020Kb

Load more
Recommended publications
-

Nanoparticles to Upregulate Notch Signaling Victor
Notch Intracellular Domain Plasmid Delivery via Poly(lactic-co-glycolic acid) Nanoparticles to Upregulate Notch Signaling Victoria L. Messerschmidt1,2†, Aneetta E. Kuriakose1,2†, Uday Chintapula1, Samantha Laboy1, Thuy Thi Dang Truong1, LeNaiya A. Kydd1, Justyn Jaworski1, Kytai T. Nguyen1,2*, Juhyun Lee1,2* 1Department of Bioengineering, University of Texas at Arlington, Arlington TX 76010 USA 2University of Texas Southwestern Medical Center, Dallas TX 75390 USA † These authors have contributed equally to this work Corresponding Author: Juhyun Lee, Ph.D. Joint Department of Bioengineering UT Arlington / UT Southwestern Arlington, TX 75022 Email: [email protected] Telephone: 817-272-6534 Fax: 817-272-2251 Abstract Notch signaling is a highly conserved signaling system that is required for embryonic development and regeneration of organs. When the signal is lost, maldevelopment occurs and leads to a lethal state. Liposomes and retroviruses are most commonly used to deliver genetic material to cells. However, there are many drawbacks to these systems such as increased toxicity, nonspecific delivery, short half-life, and stability after formulation. We utilized the negatively charged and FDA approved polymer poly(lactic-co-glycolic acid) to encapsulate Notch Intracellular Domain- containing plasmid in nanoparticles. In this study, we show that primary human umbilical vein endothelial cells readily uptake the nanoparticles with and without specific antibody targets. We demonstrated that our nanoparticles also are nontoxic, stable over time, and compatible with blood. We also determined that we can successfully transfect primary human umbilical vein endothelial cells (HUVECs) with our nanoparticles in static and dynamic environments. Lastly, we elucidated that our transfection upregulates the downstream genes of Notch signaling, indicating that the payload was viable and successfully altered the genetic downstream effects. -

Lncegfl7os Regulates Human Angiogenesis by Interacting
RESEARCH ARTICLE LncEGFL7OS regulates human angiogenesis by interacting with MAX at the EGFL7/miR-126 locus Qinbo Zhou1†, Bo Yu1†*, Chastain Anderson1, Zhan-Peng Huang2, Jakub Hanus1, Wensheng Zhang3, Yu Han4, Partha S Bhattacharjee5, Sathish Srinivasan6, Kun Zhang3, Da-zhi Wang2, Shusheng Wang1,7* 1Department of Cell and Molecular Biology, Tulane University, New Orleans, United States; 2Department of Cardiology, Boston Children’s Hospital, Harvard Medical School, Boston, United States; 3Department of Computer Science, Xavier University, New Orleans, United States; 4Aab Cardiovascular Research Institute, University of Rochester School of Medicine and Dentistry, Rochester, United States; 5Department of Biology, Xavier University, New Orleans, United States; 6Cardiovascular Biology Research Program, Oklahoma Medical Research Foundation, Oklahoma, United States; 7Department of Ophthalmology, Tulane University, New Orleans, United States Abstract In an effort to identify human endothelial cell (EC)-enriched lncRNAs,~500 lncRNAs were shown to be highly restricted in primary human ECs. Among them, lncEGFL7OS, located in the opposite strand of the EGFL7/miR-126 gene, is regulated by ETS factors through a bidirectional promoter in ECs. It is enriched in highly vascularized human tissues, and upregulated in the hearts of dilated cardiomyopathy patients. LncEGFL7OS silencing impairs angiogenesis as shown by EC/fibroblast co-culture, in vitro/in vivo and ex vivo human choroid sprouting angiogenesis assays, while lncEGFL7OS overexpression has the opposite function. Mechanistically, *For correspondence: lncEGFL7OS is required for MAPK and AKT pathway activation by regulating EGFL7/miR-126 [email protected] (BY); expression. MAX protein was identified as a lncEGFL7OS-interacting protein that functions to [email protected] (SW) regulate histone acetylation in the EGFL7/miR-126 promoter/enhancer. -

Split Deoxyribozyme Probe for Efficient Detection of Highly Structured RNA Targets
University of Central Florida STARS Honors Undergraduate Theses UCF Theses and Dissertations 2018 Split Deoxyribozyme Probe For Efficient Detection of Highly Structured RNA Targets Sheila Raquel Solarez University of Central Florida Part of the Biochemistry Commons, and the Biology Commons Find similar works at: https://stars.library.ucf.edu/honorstheses University of Central Florida Libraries http://library.ucf.edu This Open Access is brought to you for free and open access by the UCF Theses and Dissertations at STARS. It has been accepted for inclusion in Honors Undergraduate Theses by an authorized administrator of STARS. For more information, please contact [email protected]. Recommended Citation Solarez, Sheila Raquel, "Split Deoxyribozyme Probe For Efficient Detection of Highly Structured RNA Targets" (2018). Honors Undergraduate Theses. 311. https://stars.library.ucf.edu/honorstheses/311 SPLIT DEOXYRIBOZYME PROBE FOR EFFICIENT DETECTION OF HIGHLY STRUCTURED RNA TARGETS By SHEILA SOLAREZ A thesis submitted in partial fulfillment of the requirements for the Honors in the Major Program in Biological Sciences in the College of Sciences and the Burnett Honors College at the University of Central Florida Orlando, Florida Spring Term, 2018 Thesis Chair: Yulia Gerasimova, PhD ABSTRACT Transfer RNAs (tRNAs) are known for their role as adaptors during translation of the genetic information and as regulators for gene expression; uncharged tRNAs regulate global gene expression in response to changes in amino acid pools in the cell. Aminoacylated tRNAs play a role in non-ribosomal peptide bond formation, post-translational protein labeling, modification of phospholipids in the cell membrane, and antibiotic biosynthesis. [1] tRNAs have a highly stable structure that can present a challenge for their detection using conventional techniques. -

Mrna Vaccine Era—Mechanisms, Drug Platform and Clinical Prospection
International Journal of Molecular Sciences Review mRNA Vaccine Era—Mechanisms, Drug Platform and Clinical Prospection 1, 1, 2 1,3, Shuqin Xu y, Kunpeng Yang y, Rose Li and Lu Zhang * 1 State Key Laboratory of Genetic Engineering, Institute of Genetics, School of Life Science, Fudan University, Shanghai 200438, China; [email protected] (S.X.); [email protected] (K.Y.) 2 M.B.B.S., School of Basic Medical Sciences, Peking University Health Science Center, Beijing 100191, China; [email protected] 3 Shanghai Engineering Research Center of Industrial Microorganisms, Shanghai 200438, China * Correspondence: [email protected]; Tel.: +86-13524278762 These authors contributed equally to this work. y Received: 30 July 2020; Accepted: 30 August 2020; Published: 9 September 2020 Abstract: Messenger ribonucleic acid (mRNA)-based drugs, notably mRNA vaccines, have been widely proven as a promising treatment strategy in immune therapeutics. The extraordinary advantages associated with mRNA vaccines, including their high efficacy, a relatively low severity of side effects, and low attainment costs, have enabled them to become prevalent in pre-clinical and clinical trials against various infectious diseases and cancers. Recent technological advancements have alleviated some issues that hinder mRNA vaccine development, such as low efficiency that exist in both gene translation and in vivo deliveries. mRNA immunogenicity can also be greatly adjusted as a result of upgraded technologies. In this review, we have summarized details regarding the optimization of mRNA vaccines, and the underlying biological mechanisms of this form of vaccines. Applications of mRNA vaccines in some infectious diseases and cancers are introduced. It also includes our prospections for mRNA vaccine applications in diseases caused by bacterial pathogens, such as tuberculosis. -

Targetome Analysis Revealed Involvement of Mir-126 in Neurotrophin Signaling Pathway: a Possible Role in Prevention of Glioma Development
Original Article Targetome Analysis Revealed Involvement of MiR-126 in Neurotrophin Signaling Pathway: A Possible Role in Prevention of Glioma Development 1# 1, 2# 3 4, 5 Maedeh Rouigari, M.Sc. , Moein Dehbashi, Ph.D. , Kamran Ghaedi, Ph.D. , Meraj Pourhossein, Ph.D. * 1. Isfahan Neuroscience Research Center (INRC), Alzahra Hospital, Isfahan University of Medical Sciences, Isfahan, Iran 2. Genetics Division, Department of Biology, Faculty of Sciences, University of Isfahan, Isfahan, Iran 3. Cell and Molecular Biology Division, Department of Biology, Faculty of Sciences, University of Isfahan, Isfahan, Iran 4. Department of Genetics and Molecular Biology, School of Medicine, Isfahan University of Medical Sciences Isfahan, Iran 5. Department of Food Science and Technology, Food Security Research Center, School of Nutrition and Food Science, Isfahan, Iran #The first two authors equally contributed to this article. *Corresponding Address: P.O.Box: 81746-73461, Hezar Jarib Street, Department of Genetics and Molecular Biology, School of Medicine, Isfahan University of Medical Sciences, Isfahan, Iran Email: [email protected] Received: 21/Nov/2016, Accepted: 14/Mar/2017 Abstract Objective: For the first time, we used molecular signaling pathway enrichment analysis to determine possible involvement of miR-126 and IRS-1 in neurotrophin pathway. Materials and Methods: In this prospective study, validated and predicted targets (targetome) of miR-126 were collected following searching miRtarbase (http://mirtarbase.mbc.nctu.edu.tw/) and miRWalk 2.0 databases, respectively. Then, approximate expression of miR-126 targeting in Glioma tissue was examined using UniGene database (http://www.ncbi. nlm.nih.gov/unigene). In silico molecular pathway enrichment analysis was carried out by DAVID 6.7 database (http://david. -

Expanding the Genetic Code Lei Wang and Peter G
Reviews P. G. Schultz and L. Wang Protein Science Expanding the Genetic Code Lei Wang and Peter G. Schultz* Keywords: amino acids · genetic code · protein chemistry Angewandte Chemie 34 2005 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim DOI: 10.1002/anie.200460627 Angew. Chem. Int. Ed. 2005, 44,34–66 Angewandte Protein Science Chemie Although chemists can synthesize virtually any small organic molecule, our From the Contents ability to rationally manipulate the structures of proteins is quite limited, despite their involvement in virtually every life process. For most proteins, 1. Introduction 35 modifications are largely restricted to substitutions among the common 20 2. Chemical Approaches 35 amino acids. Herein we describe recent advances that make it possible to add new building blocks to the genetic codes of both prokaryotic and 3. In Vitro Biosynthetic eukaryotic organisms. Over 30 novel amino acids have been genetically Approaches to Protein encoded in response to unique triplet and quadruplet codons including Mutagenesis 39 fluorescent, photoreactive, and redox-active amino acids, glycosylated 4. In Vivo Protein amino acids, and amino acids with keto, azido, acetylenic, and heavy-atom- Mutagenesis 43 containing side chains. By removing the limitations imposed by the existing 20 amino acid code, it should be possible to generate proteins and perhaps 5. An Expanded Code 46 entire organisms with new or enhanced properties. 6. Outlook 61 1. Introduction The genetic codes of all known organisms specify the same functional roles to amino acid residues in proteins. Selectivity 20 amino acid building blocks. These building blocks contain a depends on the number and reactivity (dependent on both limited number of functional groups including carboxylic steric and electronic factors) of a particular amino acid side acids and amides, a thiol and thiol ether, alcohols, basic chain. -
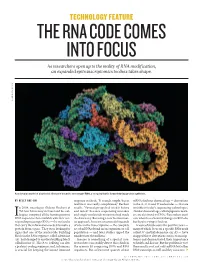
THE RNA CODE COMES INTO FOCUS As Researchers Open up to the Reality of RNA Modification, an Expanded Epitranscriptomics Toolbox Takes Shape
TECHNOLOGY FEATURE THE RNA CODE COMES INTO FOCUS As researchers open up to the reality of RNA modification, an expanded epitranscriptomics toolbox takes shape. LAGUNA DESIGN/SPL LAGUNA A molecular model of a bacterial ribosome bound to messenger RNA, a complex that is formed during protein synthesis. BY KELLY RAE CHI response in check. “It sounds simple, but in mRNAs harbour chemical tags — decorations real life it was really complicated,” Rechavi to the A, C, G and U nucleotides — that are n 2004, oncologist Gideon Rechavi at recalls. “Several groups had tried it before invisible to today’s sequencing technologies. Tel Aviv University in Israel and his col- and failed” because sequencing mistakes (Similar chemical tags, called epigenetic mark- leagues compared all the human genomic and single-nucleotide mutations had made ers, are also found on DNA.) Researchers aren’t IDNA sequences then available with their cor- the data noisy. But using a new bioinformat- sure what these chemical changes in RNA do, responding messenger RNAs — the molecules ics approach, his team uncovered thousands but they’re trying to find out. that carry the information needed to make a of sites in the transcriptome — the complete A wave of studies over the past five years — protein from a gene. They were looking for set of mRNAs found in an organism or cell many of which focus on a specific RNA mark signs that one of the nucleotide building population — and later studies upped the called N6-methyladenosine (m6A) — have blocks in the RNA sequence, called adenosine number into the millions1. -

Mrna, Rrna and Trna Types of RNA: Mrna, Rrna and Trna
Types of RNA: mRNA, rRNA and tRNA Types of RNA: mRNA, rRNA and tRNA By Susha Cheriyedath, M.Sc. Reviewed by Michael Greenwood, M.Sc. RNA or ribonucleic acid is a polymer of nucleotides that is made up of a ribose sugar, a phosphate, and bases such as adenine, guanine, cytosine, and uracil. It plays a crucial role in gene expression by acting as the intermediate between the genetic information encoded by DNA and proteins. Designua | Shutterstock RNA has a structure very similar to that of DNA. The key difference in RNA structure is that the ribose sugar in RNA possesses a hydroxyl (OH) group that is absent in DNA. Types of RNA In both prokaryotes and eukaryotes, there are three main types of RNA – messenger RNA (mRNA), ribosomal RNA (rRNA), and transfer RNA (tRNA). These 3 types of RNA are discussed below. P Saved from URL: https://www.news-medical.net/life-sciences/-Types-of-RNA-mRNA-rRNA-and-tRNA.aspx 1/5 Types of RNA: mRNA, rRNA and tRNA Messenger RNA (mRNA) mRNA accounts for just 5% of the total RNA in the cell. mRNA is the most heterogeneous of the 3 types of RNA in terms of both base sequence and size. It carries complimentary genetic code copied, from DNA during transcription, in the form of triplets of nucleotides called codons. Each codon specifies a particular amino acid, though one amino acid may be coded for by many different codons. Although there are 64 possible codons or triplet bases in the genetic code, only 20 of them represent amino acids. -

Transcription Study Guide This Study Guide Is a Written Version of the Material You Have Seen Presented in the Transcription Unit
Transcription Study Guide This study guide is a written version of the material you have seen presented in the transcription unit. The cell’s DNA contains the instructions for carrying out the work of the cell. These instructions are used by the cell’s protein-making machinery to create proteins. If the cell’s DNA were directly read by the protein-making machinery, however, it could be damaged and the process would be slow and cumbersome. The cell avoids this problem by copying genetic information from its DNA into an intermediate called messenger RNA (mRNA). It is this mRNA that is read by the cell’s protein-making machinery. This process is called transcription. Components In this section you will be introduced to the components involved in the process of RNA synthesis, called transcription. This process requires an enzyme that uses many nucleotide bases to copy the instructions present in DNA into an intermediate messenger RNA molecule. RNA What is RNA? · Like DNA, RNA is a polymer made up of nucleotides. · Unlike DNA, which is composed of two strands of nucleotides twisted together, RNA is single-stranded. It can also sometimes fold into complex three-dimensional structures. · RNA contains the same nucleotides as DNA, with the substitution of uraciluridine (U) for thymidine (T). · RNA is chemically different from DNA so that the cell can easily tell the two apart. · In this animation, you will see one type of RNA, messenger RNA, being put together. · There are three types of RNA: mRNA, which you will read more about; tRNA, which is used in the translation process, and rRNA, which acts as a structural element in the ribosome (a translation component). -

Isolation of RNA with Properties of Messenger RNA from Cerebral Polyribosomes* Claire E
Proceedings of the National Academy of Sciences Vol. 67, No. 2, pp. 644-651, October 1970 Isolation of RNA with Properties of Messenger RNA from Cerebral Polyribosomes* Claire E. Zomzelyt, Sidney Roberts, and Susan Peache DEPARTMENT OF BIOLOGICAL CHEMISTRY, SCHOOL OF MEDICINE, AND THE BRAIN RESEARCH INSTI- TUTE, UNIVERSITY OF CALIFORNIA CENTER FOR THE HEALTH SCIENCES, LOS ANGELES, CALIFORNIA 90024 Communicated by H. W. Magoun, July 24, 1970 Abstract. RNA which dissociated from purified cerebral polyribosomes of adult rats in the presence of EDTA was isolated by fractionation in a dis- continuous sucrose gradient. The yield was 2% of the total polyribosomal RNA. The base composition resembled the complementary values for rat DNA and was very different from base compositions of ribosomal RNA and transfer RNA. This RNA fraction contained a large proportion of molecules which were rapidly labeled in vivo and hybridized to homologous DNA. The polyribosomal RNA preparation also exhibited high template activity in a cerebral cell-free system which had previously been stripped of the capacity to incorporate amino acids in the absence of added messenger RNA (mRNA). Sedimentation analysis revealed only two peaks, with coefficients of approximately 8 S and 16 S. The data indicate that RNA with the properties of mRNA can be selectively isolated from cerebral polyribosomes under mild conditions which avoid degradation. Numerous studies support the concept that alterations in RNA and protein synthesis are associated with information transfer and storage in the central nervous system. Activation of the synthesis of specific proteins is suggested by the reports that new RNA with altered base composition, presumably mRNA, is formed during the acquisition phase of learning. -

Why We Should Take a Second Look at RNA Technology
RNA THERAPEUTICS RNA Modulation: Gabriele Campi Why we should take a second look at RNA technology As a scientist first, but also as an investor, I have always hereditary ATTR amyloidosis with polyneuropathy, an believed in the potential of RNA molecules as therapeutics. autosomal dominant neurodegenerative disease. In 2018 While most current drugs aim to stop disease by modulating patisiran (Onpattro) was approved by the FDA. It is existing proteins, RNA molecules operate one step earlier. delivered to cells by lipid nanoparticles rather than GalNAc In the case of RNA interference (RNAi), they modulate the (N-Acetylgalactoseamine-siRNA conjugates). genes involved in different pathological processes. The RNA Since the end of 2018, investors have made a cautious molecules, small interfering RNA and microRNA, both return to the RNA therapy space with a targeted funding degrade messenger RNA (mRNA) and prevent it from being of existing RNA companies. In parallel, new companies are translated into proteins. entering the arena. An unexpected boost to the industry has I am the co-founder of the investment company AurorA come from the development and approval of the first mRNA Science, which recently led a Series B financing round for the vaccines to treat Covid-19 from Pfizer/BioNTech and Moderna. Dutch company InteRNA Technologies, which is developing I have always believed in the potency of RNA interference RNA therapeutics for the treatment of advanced solid and modulation techniques that allow a researcher to target tumours. This is an ambitious project, but with a potential specific messenger RNAs through the use of siRNAs, or for success. -

Pfizer-Biontech COVID-19 Vaccine)
Nucleoside-modified messenger RNA (modmRNA) of SARS-CoV-2 Suspension for injection (Pfizer-BioNTech COVID-19 Vaccine) U.S. Trade Names Pfizer-BioNTech COVID-19 The list of names may not include all products that are available on the market. What is this medicine? SARS COVID-19 VACCINE (SARZ koh-vid 19 vak SEEN) is a vaccine used to reduce the risk of getting COVID-19. This vaccine does not treat COVID-19. There is no FDA-approved vaccine to prevent COVID-19. The FDA has authorized the emergency use of this vaccine during the COVID-19 pandemic. This medicine may be used for other purposes; ask your health care provider or pharmacist if you have questions. What should I tell my health care provider before I take this medicine? They need to know if you have any of these conditions: any allergies bleeding disorder fever or infection immune system problems recent or upcoming vaccine including previous COVID-19 vaccine an unusual or allergic reaction to COVID-19 vaccine, other drugs, foods, dyes, or preservatives pregnant or trying to get pregnant breast-feeding How should I use this medicine? This vaccine is injected into a muscle. It is given by a health care professional. A copy of the Fact Sheet for Recipients and Caregivers will be given before each vaccination. Be sure to read this information carefully each time. This sheet may change frequently. Talk to your health care provider about the use of this drug in children. Special care may be needed. Overdosage: If you think you have taken too much of this medicine contact a poison control center or emergency room at once.