Inhibition of Riboflavin Metabolism in Rat Tissues by Chlorpromazine, Imipramine, and Amitriptyline
Total Page:16
File Type:pdf, Size:1020Kb

Load more
Recommended publications
-

First Xenon-Xenon Collisions in the Lhc
9th International Particle Accelerator Conference IPAC2018, Vancouver, BC, Canada JACoW Publishing ISBN: 978-3-95450-184-7 doi:10.18429/JACoW-IPAC2018-MOPMF039 FIRST XENON-XENON COLLISIONS IN THE LHC M. Schaumann∗, R. Alemany-Fernandez, P. Baudrenghien, T. Bohl, C. Bracco, R. Bruce, N. Fuster-Martinez, M. A. Jebramcik, J.M. Jowett, T. Mertens, D. Mirarchi, S. Redaelli, B. Salvachua, M. Solfaroli, H. Timko, J. Wenninger, CERN, Geneva, Switzerland Abstract EXECUTION OF THE RUN In 2017, the CERN accelerator complex once again demonstrated its flexibility by producing beams of a new A total of about 18 h of LHC time were taken for the Xe ion species, xenon, that were successfully injected into LHC. collision run. The schedule was designed to include set-up, On 12 October, collisions of fully stripped xenon nuclei first injection, validation and physics data taking. A further 12 h were devoted to experiments on crystal collimation, de- were recorded for the first time in the LHC√ at a centre-of- scribed elsewhere [3]. This tight schedule was only feasible mass energy per colliding nucleon pair of sNN = 5.44 TeV. Physics data taking started 9.5hafter the first injection of by drastically reducing the complexity of the set-up follow- xenon beams and lasted a total of 6h. The integrated lumi- ing the model of the p–Pb pilot run in September 2012 [4,5] nosity delivered to the four LHC experiments was sufficient when first injection, validation and physics data-taking were that new physics results can be expected soon. We provide achieved within a single fill. -

XENON X-1100 High-Intensity Pulsed Light System
XENON X-1100 High-Intensity Pulsed Light System The low cost R&D tool for investigating the applicability of Pulsed Light for new and emerging applications. The tool that researchers, scientists and engineers have been looking for is here. An easy-to-use photonic system developed by XENON that will open new doors and lead to new discoveries. The power of Pulsed Light In virtually all disciplines of science and technology there are applications where precise delivery of energy is demanded. One less studied energy delivery mechanism is the use of high-intensity pulsed light. At present the most prevalent example of this is in the use of lasers. However, the point source, coherent and single wavelength nature of lasers, are often not suited to the majority of challenges facing many industries. In these situations, it is crucial to have a broad spectra source so as not to be restricted in choosing materials with specific absorption characteristics. Because XENON sources generate light which extends from the deep UV to IR and often with peak pulse powers measured in megawatts, the ability of these sources to perform tasks such as breaking chemical bonds, sintering, ablating or sterilizing is highly realizable. The high peak powers generated by XENON’s production level systems are possible by tightly controlling the pulse durations measured from a few microseconds to milliseconds. Until now there was no practical method of generating this intense pulsed light in a low-cost R&D tool that was safe, compact and versatile. XENON has developed a system to address this challenge based on over 50 years of exper- tise in the Pulsed Light industry. -
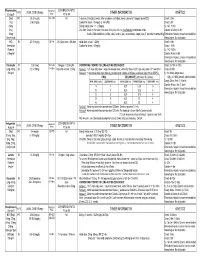
Medication Conversion Chart
Fluphenazine FREQUENCY CONVERSION RATIO ROUTE USUAL DOSE (Range) (Range) OTHER INFORMATION KINETICS Prolixin® PO to IM Oral PO 2.5-20 mg/dy QD - QID NA ↑ dose by 2.5mg/dy Q week. After symptoms controlled, slowly ↓ dose to 1-5mg/dy (dosed QD) Onset: ≤ 1hr 1mg (2-60 mg/dy) Caution for doses > 20mg/dy (↑ risk EPS) Cmax: 0.5hr 2.5mg Elderly: Initial dose = 1 - 2.5mg/dy t½: 14.7-15.3hr 5mg Oral Soln: Dilute in 2oz water, tomato or fruit juice, milk, or uncaffeinated carbonated drinks Duration of Action: 6-8hr 10mg Avoid caffeinated drinks (coffee, cola), tannics (tea), or pectinates (apple juice) 2° possible incompatibilityElimination: Hepatic to inactive metabolites 5mg/ml soln Hemodialysis: Not dialyzable HCl IM 2.5-10 mg/dy Q6-8 hr 1/3-1/2 po dose = IM dose Initial dose (usual): 1.25mg Onset: ≤ 1hr Immediate Caution for doses > 10mg/dy Cmax: 1.5-2hr Release t½: 14.7-15.3hr 2.5mg/ml Duration Action: 6-8hr Elimination: Hepatic to inactive metabolites Hemodialysis: Not dialyzable Decanoate IM 12.5-50mg Q2-3 wks 10mg po = 12.5mg IM CONVERTING FROM PO TO LONG-ACTING DECANOATE: Onset: 24-72hr (4-72hr) Long-Acting SC (12.5-100mg) (1-4 wks) Round to nearest 12.5mg Method 1: 1.25 X po daily dose = equiv decanoate dose; admin Q2-3wks. Cont ½ po daily dose X 1st few mths Cmax: 48-96hr 25mg/ml Method 2: ↑ decanoate dose over 4wks & ↓ po dose over 4-8wks as follows (accelerate taper for sx of EPS): t½: 6.8-9.6dy (single dose) ORAL DECANOATE (Administer Q 2 weeks) 15dy (14-100dy chronic administration) ORAL DOSE (mg/dy) ↓ DOSE OVER (wks) INITIAL DOSE (mg) TARGET DOSE (mg) DOSE OVER (wks) Steady State: 2mth (1.5-3mth) 5 4 6.25 6.25 0 Duration Action: 2wk (1-6wk) Elimination: Hepatic to inactive metabolites 10 4 6.25 12.5 4 Hemodialysis: Not dialyzable 20 8 6.25 12.5 4 30 8 6.25 25 4 40 8 6.25 25 4 Method 3: Admin equivalent decanoate dose Q2-3wks. -

Adverse Reactions to Hallucinogenic Drugs. 1Rnstttutton National Test
DOCUMENT RESUME ED 034 696 SE 007 743 AUTROP Meyer, Roger E. , Fd. TITLE Adverse Reactions to Hallucinogenic Drugs. 1rNSTTTUTTON National Test. of Mental Health (DHEW), Bethesda, Md. PUB DATP Sep 67 NOTE 118p.; Conference held at the National Institute of Mental Health, Chevy Chase, Maryland, September 29, 1967 AVATLABLE FROM Superintendent of Documents, Government Printing Office, Washington, D. C. 20402 ($1.25). FDPS PRICE FDPS Price MFc0.50 HC Not Available from EDRS. DESCPTPTOPS Conference Reports, *Drug Abuse, Health Education, *Lysergic Acid Diethylamide, *Medical Research, *Mental Health IDENTIFIEPS Hallucinogenic Drugs ABSTPACT This reports a conference of psychologists, psychiatrists, geneticists and others concerned with the biological and psychological effects of lysergic acid diethylamide and other hallucinogenic drugs. Clinical data are presented on adverse drug reactions. The difficulty of determining the causes of adverse reactions is discussed, as are different methods of therapy. Data are also presented on the psychological and physiolcgical effects of L.S.D. given as a treatment under controlled medical conditions. Possible genetic effects of L.S.D. and other drugs are discussed on the basis of data from laboratory animals and humans. Also discussed are needs for futher research. The necessity to aviod scare techniques in disseminating information about drugs is emphasized. An aprentlix includes seven background papers reprinted from professional journals, and a bibliography of current articles on the possible genetic effects of drugs. (EB) National Clearinghouse for Mental Health Information VA-w. Alb alb !bAm I.S. MOMS Of NAM MON tMAN IONE Of NMI 105 NUNN NU IN WINES UAWAS RCM NIN 01 NUN N ONMININI 01011110 0. -

Pharmacology – Inhalant Anesthetics
Pharmacology- Inhalant Anesthetics Lyon Lee DVM PhD DACVA Introduction • Maintenance of general anesthesia is primarily carried out using inhalation anesthetics, although intravenous anesthetics may be used for short procedures. • Inhalation anesthetics provide quicker changes of anesthetic depth than injectable anesthetics, and reversal of central nervous depression is more readily achieved, explaining for its popularity in prolonged anesthesia (less risk of overdosing, less accumulation and quicker recovery) (see table 1) Table 1. Comparison of inhalant and injectable anesthetics Inhalant Technique Injectable Technique Expensive Equipment Cheap (needles, syringes) Patent Airway and high O2 Not necessarily Better control of anesthetic depth Once given, suffer the consequences Ease of elimination (ventilation) Only through metabolism & Excretion Pollution No • Commonly administered inhalant anesthetics include volatile liquids such as isoflurane, halothane, sevoflurane and desflurane, and inorganic gas, nitrous oxide (N2O). Except N2O, these volatile anesthetics are chemically ‘halogenated hydrocarbons’ and all are closely related. • Physical characteristics of volatile anesthetics govern their clinical effects and practicality associated with their use. Table 2. Physical characteristics of some volatile anesthetic agents. (MAC is for man) Name partition coefficient. boiling point MAC % blood /gas oil/gas (deg=C) Nitrous oxide 0.47 1.4 -89 105 Cyclopropane 0.55 11.5 -34 9.2 Halothane 2.4 220 50.2 0.75 Methoxyflurane 11.0 950 104.7 0.2 Enflurane 1.9 98 56.5 1.68 Isoflurane 1.4 97 48.5 1.15 Sevoflurane 0.6 53 58.5 2.5 Desflurane 0.42 18.7 25 5.72 Diethyl ether 12 65 34.6 1.92 Chloroform 8 400 61.2 0.77 Trichloroethylene 9 714 86.7 0.23 • The volatile anesthetics are administered as vapors after their evaporization in devices known as vaporizers. -

Neuroprotection by Chlorpromazine and Promethazine in Severe Transient and Permanent Ischemic Stroke
Mol Neurobiol (2017) 54:8140–8150 DOI 10.1007/s12035-016-0280-x Neuroprotection by Chlorpromazine and Promethazine in Severe Transient and Permanent Ischemic Stroke Xiaokun Geng1,2 & Fengwu Li1 & James Yip2 & Changya Peng2 & Omar Elmadhoun2 & Jiamei Shen1 & Xunming Ji1,3 & Yuchuan Ding1,2 Received: 29 June 2016 /Accepted: 31 October 2016 /Published online: 28 November 2016 # Springer Science+Business Media New York 2016 Abstract Previous studies have demonstrated depressive or enhance C + P-induced neuroprotection. C + P therapy im- hibernation-like roles of phenothiazine neuroleptics [com- proved brain metabolism as determined by increased ATP bined chlorpromazine and promethazine (C + P)] in brain levels and NADH activity, as well as decreased ROS produc- activity. This ischemic stroke study aimed to establish neuro- tion. These therapeutic effects were associated with alterations protection by reducing oxidative stress and improving brain in PKC-δ and Akt protein expression. C + P treatments con- metabolism with post-ischemic C + P administration. ferred neuroprotection in severe stroke models by suppressing Sprague-Dawley rats were subjected to transient (2 or 4 h) the damaging cascade of metabolic events, most likely inde- middle cerebral artery occlusion (MCAO) followed by 6 or pendent of drug-induced hypothermia. These findings further 24 h reperfusion, or permanent (28 h) MCAO without reper- prove the clinical potential for C + P treatment and may direct fusion. At 2 h after ischemia onset, rats received either an us closer towards the development of an efficacious neuropro- intraperitoneal (IP) injection of saline or two doses of C + P. tective therapy. Body temperatures, brain infarct volumes, and neurological deficits were examined. -

Is Aristada (Aripiprazole Lauroxil) a Safe and Effective Treatment for Schizophrenia in Adult Patients? Kyle J
Philadelphia College of Osteopathic Medicine DigitalCommons@PCOM PCOM Physician Assistant Studies Student Student Dissertations, Theses and Papers Scholarship 2017 Is Aristada (Aripiprazole Lauroxil) a Safe and Effective Treatment For Schizophrenia In Adult Patients? Kyle J. Knowles Philadelphia College of Osteopathic Medicine Follow this and additional works at: https://digitalcommons.pcom.edu/pa_systematic_reviews Part of the Psychiatry Commons Recommended Citation Knowles, Kyle J., "Is Aristada (Aripiprazole Lauroxil) a Safe and Effective Treatment For Schizophrenia In Adult Patients?" (2017). PCOM Physician Assistant Studies Student Scholarship. 381. https://digitalcommons.pcom.edu/pa_systematic_reviews/381 This Selective Evidence-Based Medicine Review is brought to you for free and open access by the Student Dissertations, Theses and Papers at DigitalCommons@PCOM. It has been accepted for inclusion in PCOM Physician Assistant Studies Student Scholarship by an authorized administrator of DigitalCommons@PCOM. For more information, please contact [email protected]. Is Aristada (Aripiprazole Lauroxil) a Safe and Effective Treatment For Schizophrenia In Adult Patients? Kyle J. Knowles, PA-S A SELECTIVE EVIDENCE BASED MEDICINE REVIEW In Partial Fulfillment of the Requirements For The Degree of Master of Science In Health Sciences- Physician Assistant Department of Physician Assistant Studies Philadelphia College of Osteopathic Medicine Philadelphia, Pennsylvania December 16, 2016 ABSTRACT OBJECTIVE: The objective of this selective EBM review is to determine whether or not “Is Aristada (aripiprazole lauroxil) a safe and effective treatment for schizophrenia in adult patients?” STUDY DESIGN: Review of three randomized controlled studies. All three trials were conducted between 2014 and 2015. DATA SOURCES: One randomized, controlled trial and two randomized, controlled, double- blind trials found via Cochrane Library and PubMed. -

Xenon Gas Xe Safety Data Sheet SDS P4677
Xenon Safety Data Sheet P-4677 This SDS conforms to U.S. Code of Federal Regulations 29 CFR 1910.1200, Hazard Communication. Date of issue: 01/01/1979 Revision date: 10/24/2016 Supersedes: 10/02/2014 SECTION: 1. Product and company identification 1.1. Product identifier Product form : Substance Name : Xenon CAS No : 7440-63-3 Formula : Xe Other means of identification : Xenon 1.2. Relevant identified uses of the substance or mixture and uses advised against Use of the substance/mixture : Industrial use. Use as directed. 1.3. Details of the supplier of the safety data sheet Praxair, Inc. 10 Riverview Drive Danbury, CT 06810-6268 - USA T 1-800-772-9247 (1-800-PRAXAIR) - F 1-716-879-2146 www.praxair.com 1.4. Emergency telephone number Emergency number : Onsite Emergency: 1-800-645-4633 CHEMTREC, 24hr/day 7days/week — Within USA: 1-800-424-9300, Outside USA: 001-703-527-3887 (collect calls accepted, Contract 17729) SECTION 2: Hazard identification 2.1. Classification of the substance or mixture GHS-US classification Compressed gas H280 2.2. Label elements GHS-US labeling Hazard pictograms (GHS-US) : GHS04 Signal word (GHS-US) : WARNING Hazard statements (GHS-US) : H280 - CONTAINS GAS UNDER PRESSURE; MAY EXPLODE IF HEATED OSHA-H01 - MAY DISPLACE OXYGEN AND CAUSE RAPID SUFFOCATION Precautionary statements (GHS-US) : P202 - Do not handle until all safety precautions have been read and understood P271+P403 - Use and store only outdoors or in a well-ventilated place CGA-PG05 - Use a back flow preventive device in the piping CGA-PG10 - Use only with equipment rated for cylinder pressure CGA-PG06 - Close valve after each use and when empty CGA-PG02 - Protect from sunlight when ambient temperature exceeds 52°C (125°F) 2.3. -

Antipsychotic Combinations
Graylands Hospital DRUG BULLETIN Pharmacy Department Brockway Road Mount Claremont WA 6010 Telephone (08) 9347 6400 Email [email protected] Fax (08) 9384 4586 Antipsychotic Combinations Graylands Hospital Drug Bulletin 2008 Vol. 15 No. 3 February ISSN 1323-1251 Antipsychotic combinations Reasons for antipsychotic combinations Despite the development of efficacious medications for the treatment of schizophrenia, many people do There are a number of theoretical benefits and not respond adequately. To address this problem, reasons cited for antipsychotic combination the use of two or more antipsychotics simultaneously prescribing, these include: is a commonly employed treatment strategy. Although the use of combination antipsychotics is Complementary mechanisms of action4 (e.g. common in clinical practice, the risks and benefits adding an antipsychotic with strong dopamine have not been systematically evaluated to date. As a D2 blockade to a weak D2 blocker) result, current Australian treatment algorithms Partial replacement of antipsychotic action for including the Royal Australian and New Zealand drugs with intolerable adverse effects at higher College of Psychiatry Schizophrenia Guidelines and 5 doses (e.g. adding quetiapine to clozapine to the Western Australian Therapeutic Advisory Group minimise metabolic adverse effects) Antipsychotic Guidelines advise against the use of combined antipsychotics, except for short periods of Alternative where clozapine cannot be used6 changeover1,2. Most data on antipsychotic -

Lung Ventilation - Xenon
Lung Ventilation - Xenon Special Instructions Patients with severe dyspnea may not be able to tolerate this examination. To be performed only at UNMH. Radiopharmaceutical: Xe-133 gas Dose (Adult/Pediatric): Refer to Nuclear Medicine Dose Chart Route of Administration: Inhalation from a Xe-133 gas delivery system. Patient Preparation: None. Equipment Setup: Gamma Camera: ZOOM as appropriate for small children Collimator: Orbiter: LEAP SPECT-CT/ECAM/Evo/Symbia E: High resolution Computer setup: • Static acquisition • 128 x 128 matrix • Zoom 1.0 (greater for small children)—ensure that the lungs fill a significant portion of the detector for small patients • 10 sec/image for inspiration • 30 sec/image for all other images Patient Positioning: Orbiter: • Seated upright on a stool (preferred) with the detector posterior to the patient. Supine if necessary All others: • Supine Procedure: • If the patient is able to sit upright on a stool for the examination, perform the ventilation portion of the examination on the Orbiter if available. • Before beginning the study, explain the breathing maneuvers required; the patient should rehearse these maneuvers. • Place the mask on the patient and ensure that it fits snugly. • Inspiration (Single-breath) Image: • Instruct the patient to exhale completely and then to take a deep breath as the Xe-133 gas flows in. • The patient should hold his/her breath for 10 sec (if possible) while posterior (and anterior if on a dual-headed camera) imaging is performed. Revised 2017-01 Lung Ventilation - Xenon (continued) • Equilibrium (Rebreathing/Wash-in) Images: • The patient should then breathe normally for 3 images (90 seconds). -

New Drugs Are Not Enough‑Drug Repositioning in Oncology: an Update
INTERNATIONAL JOURNAL OF ONCOLOGY 56: 651-684, 2020 New drugs are not enough‑drug repositioning in oncology: An update ROMINA GABRIELA ARMANDO, DIEGO LUIS MENGUAL GÓMEZ and DANIEL EDUARDO GOMEZ Laboratory of Molecular Oncology, Science and Technology Department, National University of Quilmes, Bernal B1876, Argentina Received August 15, 2019; Accepted December 16, 2019 DOI: 10.3892/ijo.2020.4966 Abstract. Drug repositioning refers to the concept of discov- 17. Lithium ering novel clinical benefits of drugs that are already known 18. Metformin for use treating other diseases. The advantages of this are that 19. Niclosamide several important drug characteristics are already established 20. Nitroxoline (including efficacy, pharmacokinetics, pharmacodynamics and 21. Nonsteroidal anti‑inflammatory drugs toxicity), making the process of research for a putative drug 22. Phosphodiesterase-5 inhibitors quicker and less costly. Drug repositioning in oncology has 23. Pimozide received extensive focus. The present review summarizes the 24. Propranolol most prominent examples of drug repositioning for the treat- 25. Riluzole ment of cancer, taking into consideration their primary use, 26. Statins proposed anticancer mechanisms and current development 27. Thalidomide status. 28. Valproic acid 29. Verapamil 30. Zidovudine Contents 31. Concluding remarks 1. Introduction 2. Artesunate 1. Introduction 3. Auranofin 4. Benzimidazole derivatives In previous decades, a considerable amount of work has been 5. Chloroquine conducted in search of novel oncological therapies; however, 6. Chlorpromazine cancer remains one of the leading causes of death globally. 7. Clomipramine The creation of novel drugs requires large volumes of capital, 8. Desmopressin alongside extensive experimentation and testing, comprising 9. Digoxin the pioneer identification of identifiable targets and corrobora- 10. -

PERPHENAZINE and AMITRIPTYLINE HYDROCHLORIDE- Perphenazine and Amitriptyline Hydrochloride Tablet, Film Coated Mylan Pharmaceuticals Inc
PERPHENAZINE AND AMITRIPTYLINE HYDROCHLORIDE- perphenazine and amitriptyline hydrochloride tablet, film coated Mylan Pharmaceuticals Inc. ---------- WARNING Increased Mortality in Elderly Patients with Dementia-Related Psychosis Elderly patients with dementia-related psychosis treated with antipsychotic drugs are at an increased risk of death. Analyses of seventeen placebo-controlled trials (modal duration of 10 weeks), largely in patients taking atypical antipsychotic drugs, revealed a risk of death in drug-treated patients of between 1.6 to 1.7 times the risk of death in placebo-treated patients. Over the course of a typical 10-week controlled trial, the rate of death in drug- treated patients was about 4.5%, compared to a rate of about 2.6% in the placebo group. Although the causes of death were varied, most of the deaths appeared to be either cardiovascular (e.g., heart failure, sudden death) or infectious (e.g., pneumonia) in nature. Observational studies suggest that, similar to atypical antipsychotic drugs, treatment with conventional antipsychotic drugs may increase mortality. The extent to which the findings of increased mortality in observational studies may be attributed to the antipsychotic drug as opposed to some characteristic(s) of the patients is not clear. Perphenazine and amitriptyline hydrochloride is not approved for the treatment of patients with dementia- related psychosis (see WARNINGS). Suicidality and Antidepressant Drugs Antidepressants increased the risk compared to placebo of suicidal thinking and behavior (suicidality) in children, adolescents and young adults in short-term studies of major depressive disorder (MDD) and other psychiatric disorders. Anyone considering the use of perphenazine and amitriptyline or any other antidepressant in a child, adolescent, or young adult must balance this risk with the clinical need.