New Drugs Are Not Enough‑Drug Repositioning in Oncology: an Update
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Thalidomide Celgene Pregnancy Prevention Programme Information
TCPPP Healthcare Professional Booklet Thalidomide Celgene ® Pregnancy Prevention Programme Information for Healthcare Professionals ® Prescribing or Dispensing Thalidomide Celgene Celgene representative contact details: AM Mangion Ltd, Regulatory Office, “Mangion Building”, New Street Off Valletta Road, Luqa. Phone: +356 239 76333 Fax: +356 239 76123 Email: [email protected] Medical Information queries: [email protected] 2014 © Celgene Corporation Page 1 of 19 RMP/THA/001/14-03/M TCPPP Healthcare Professional Booklet This booklet is intended for healthcare professionals involved in prescribing or dispensing Thalidomide Celgene ®, and contains information about: Preventing harm to unborn babies: If Thalidomide Celgene® is taken during pregnancy it can cause severe birth defects or death to an unborn baby. Other side effects of Thalidomide Celgene®: The most commonly observed adverse reactions associated with the use of Thalidomide Celgene® in combination with melphalan and prednisone are: neutropenia, leukopenia, constipation, somnolence, paraesthesia, peripheral neuropathy, anaemia, lymphopenia, thrombocytopenia, dizziness, dysaesthesia, tremor and peripheral oedema. Further information and recommended precautions can be found in the Thalidomide Celgene® Summary of Product Characteristics (SmPC). Thalidomide Celgene® Pregnancy Prevention Programme: This Programme is designed to make sure that unborn babies are not exposed to Thalidomide Celgene®. It will provide you with information about how to follow the programme and explain your responsibilities. This booklet will help you understand these problems and make sure you know what to do before prescribing and dispensing Thalidomide Celgene®. For your patients’ own health and safety, please read this booklet carefully. You must ensure that your patients fully understand what you have told them about Thalidomide Celgene® before starting treatment. -

Practice Parameters for the Treatment of Patients with Dominantly Inherited Colorectal Cancer
Practice Parameters For The Treatment Of Patients With Dominantly Inherited Colorectal Cancer Diseases of the Colon & Rectum 2003;46(8):1001-1012 Prepared by: The Standards Task Force The American Society of Colon and Rectal Surgeons James Church, MD; Clifford Simmang, MD; On Behalf of the Collaborative Group of the Americas on Inherited Colorectal Cancer and the Standards Committee of the American Society of Colon and Rectal Surgeons. The American Society of Colon and Rectal Surgeons is dedicated to assuring high quality patient care by advancing the science, prevention, and management of disorders and diseases of the colon, rectum, and anus. The standards committee is composed of Society members who are chosen because they have demonstrated expertise in the specialty of colon and rectal surgery. This Committee was created in order to lead international efforts in defining quality care for conditions related to the colon, rectum, and anus. This is accompanied by developing Clinical Practice Guidelines based on the best available evidence. These guidelines are inclusive, and not prescriptive. Their purpose is to provide information on which decisions can be made, rather than dictate a specific form of treatment. These guidelines are intended for the use of all practitioners, health care workers, and patients who desire information about the management of the conditions addressed by the topics covered in these guidelines. Practice Parameters for the Treatment of Patients With Dominantly Inherited Colorectal Cancer Inherited colorectal cancer includes two main syndromes in which predisposition to the disease is based on a germline mutation that may be transmitted from parent to child. -

FDA Comments on CBD in Foods
Popular Content Public Health Focus FDA and Marijuana: Questions and Answers 1. How is marijuana therapy being used by some members of the medical community? 2. Why hasn’t the FDA approved marijuana for medical uses? 3. Is marijuana safe for medical use? 4. How does FDA’s role differ from NIH and DEA’s role when it comes to the investigation of marijuana for medical use? 5. Does the FDA object to the clinical investigation of marijuana for medical use? 6. What kind of research is the FDA reviewing when it comes to the efficacy of marijuana? 7. How can patients get into expanded access program for marijuana for medical use? 8. Does the FDA have concerns about administering a cannabis product to children? 9. What is FDA’s reaction to states that are allowing marijuana to be sold for medical uses without the FDA’s approval? 10. What is the FDA’s position on state “Right to Try” bills? 11. Has the agency received any adverse event reports associated with marijuana for medical conditions? 12. Can products that contain cannabidiol be sold as dietary supplements? 13. Is it legal, in interstate commerce, to sell a food to which cannabidiol has been added? 14. In making the two previous determinations, why did FDA conclude that substantial clinical investigations have been authorized for and/or instituted about cannabidiol, and that the existence of such investigations has been made public? 15. Will FDA take enforcement action regarding cannabidiol products that are marketed as dietary supplements? What about foods to which cannabidiol has been added? 16. -

Hormonal Treatment Strategies Tailored to Non-Binary Transgender Individuals
Journal of Clinical Medicine Review Hormonal Treatment Strategies Tailored to Non-Binary Transgender Individuals Carlotta Cocchetti 1, Jiska Ristori 1, Alessia Romani 1, Mario Maggi 2 and Alessandra Daphne Fisher 1,* 1 Andrology, Women’s Endocrinology and Gender Incongruence Unit, Florence University Hospital, 50139 Florence, Italy; [email protected] (C.C); jiska.ristori@unifi.it (J.R.); [email protected] (A.R.) 2 Department of Experimental, Clinical and Biomedical Sciences, Careggi University Hospital, 50139 Florence, Italy; [email protected]fi.it * Correspondence: fi[email protected] Received: 16 April 2020; Accepted: 18 May 2020; Published: 26 May 2020 Abstract: Introduction: To date no standardized hormonal treatment protocols for non-binary transgender individuals have been described in the literature and there is a lack of data regarding their efficacy and safety. Objectives: To suggest possible treatment strategies for non-binary transgender individuals with non-standardized requests and to emphasize the importance of a personalized clinical approach. Methods: A narrative review of pertinent literature on gender-affirming hormonal treatment in transgender persons was performed using PubMed. Results: New hormonal treatment regimens outside those reported in current guidelines should be considered for non-binary transgender individuals, in order to improve psychological well-being and quality of life. In the present review we suggested the use of hormonal and non-hormonal compounds, which—based on their mechanism of action—could be used in these cases depending on clients’ requests. Conclusion: Requests for an individualized hormonal treatment in non-binary transgender individuals represent a future challenge for professionals managing transgender health care. For each case, clinicians should balance the benefits and risks of a personalized non-standardized treatment, actively involving the person in decisions regarding hormonal treatment. -

Study Protocol and Statistical Analysis Plan
Official Title: A Phase III, Multicenter, Randomized Study of Atezolizumab (Anti-PD- L1 Antibody) in Combination With Enzalutamide Versus Enzalutamide Alone in Patients With Metastatic Castration-Resistant Prostate Cancer After Failure of an Androgen Synthesis Inhibitor and Failure of, Ineligibility for, or Refusal of a Taxane Regimen NCT Number: NCT03016312 Document Date: Protocol Version 8: 14 February 2020 PROTOCOL TITLE: A PHASE III, MULTICENTER, RANDOMIZED STUDY OF ATEZOLIZUMAB (ANTIPD-L1 ANTIBODY) IN COMBINATION WITH ENZALUTAMIDE VERSUS ENZALUTAMIDE ALONE IN PATIENTS WITH METASTATIC CASTRATION-RESISTANT PROSTATE CANCER AFTER FAILURE OF AN ANDROGEN SYNTHESIS INHIBITOR AND FAILURE OF, INELIGIBILITY FOR, OR REFUSAL OF A TAXANE REGIMEN PROTOCOL NUMBER: CO39385 VERSION NUMBER: 8 EUDRACT NUMBER: 2016-003092-22 IND NUMBER: 131196 TEST PRODUCTS: Atezolizumab (RO5541267) and enzalutamide MEDICAL MONITOR: , M.D., Ph.D. SPONSOR: F. Hoffmann-La Roche Ltd DATE FINAL: Version 1: 29 September 2016 DATES AMENDED: Version 2: 7 March 2017 Version 3: 4 April 2017 Version 4: 29 June 2017 Version 5: 2 March 2018 Version 6: 23 August 2018 Version 7: 5 August 2019 Version 8: See electronic date stamp below. PROTOCOL AMENDMENT APPROVAL Date and Time (UTC) Title Approver's Name 14-Feb-2020 17:27:37 Company Signatory CONFIDENTIAL This clinical study is being sponsored globally by F. Hoffmann-La Roche Ltd of Basel, Switzerland. However, it may be implemented in individual countries by Roche’s local affiliates, including Genentech, Inc. in the United States. The information contained in this document, especially any unpublished data, is the property of F. Hoffmann-La Roche Ltd (or under its control) and therefore is provided to you in confidence as an investigator, potential investigator, or consultant, for review by you, your staff, and an applicable Ethics Committee or Institutional Review Board. -

Cancer Drug Pharmacology Table
CANCER DRUG PHARMACOLOGY TABLE Cytotoxic Chemotherapy Drugs are classified according to the BC Cancer Drug Manual Monographs, unless otherwise specified (see asterisks). Subclassifications are in brackets where applicable. Alkylating Agents have reactive groups (usually alkyl) that attach to Antimetabolites are structural analogues of naturally occurring molecules DNA or RNA, leading to interruption in synthesis of DNA, RNA, or required for DNA and RNA synthesis. When substituted for the natural body proteins. substances, they disrupt DNA and RNA synthesis. bendamustine (nitrogen mustard) azacitidine (pyrimidine analogue) busulfan (alkyl sulfonate) capecitabine (pyrimidine analogue) carboplatin (platinum) cladribine (adenosine analogue) carmustine (nitrosurea) cytarabine (pyrimidine analogue) chlorambucil (nitrogen mustard) fludarabine (purine analogue) cisplatin (platinum) fluorouracil (pyrimidine analogue) cyclophosphamide (nitrogen mustard) gemcitabine (pyrimidine analogue) dacarbazine (triazine) mercaptopurine (purine analogue) estramustine (nitrogen mustard with 17-beta-estradiol) methotrexate (folate analogue) hydroxyurea pralatrexate (folate analogue) ifosfamide (nitrogen mustard) pemetrexed (folate analogue) lomustine (nitrosurea) pentostatin (purine analogue) mechlorethamine (nitrogen mustard) raltitrexed (folate analogue) melphalan (nitrogen mustard) thioguanine (purine analogue) oxaliplatin (platinum) trifluridine-tipiracil (pyrimidine analogue/thymidine phosphorylase procarbazine (triazine) inhibitor) -
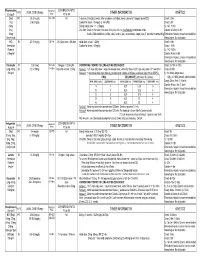
Medication Conversion Chart
Fluphenazine FREQUENCY CONVERSION RATIO ROUTE USUAL DOSE (Range) (Range) OTHER INFORMATION KINETICS Prolixin® PO to IM Oral PO 2.5-20 mg/dy QD - QID NA ↑ dose by 2.5mg/dy Q week. After symptoms controlled, slowly ↓ dose to 1-5mg/dy (dosed QD) Onset: ≤ 1hr 1mg (2-60 mg/dy) Caution for doses > 20mg/dy (↑ risk EPS) Cmax: 0.5hr 2.5mg Elderly: Initial dose = 1 - 2.5mg/dy t½: 14.7-15.3hr 5mg Oral Soln: Dilute in 2oz water, tomato or fruit juice, milk, or uncaffeinated carbonated drinks Duration of Action: 6-8hr 10mg Avoid caffeinated drinks (coffee, cola), tannics (tea), or pectinates (apple juice) 2° possible incompatibilityElimination: Hepatic to inactive metabolites 5mg/ml soln Hemodialysis: Not dialyzable HCl IM 2.5-10 mg/dy Q6-8 hr 1/3-1/2 po dose = IM dose Initial dose (usual): 1.25mg Onset: ≤ 1hr Immediate Caution for doses > 10mg/dy Cmax: 1.5-2hr Release t½: 14.7-15.3hr 2.5mg/ml Duration Action: 6-8hr Elimination: Hepatic to inactive metabolites Hemodialysis: Not dialyzable Decanoate IM 12.5-50mg Q2-3 wks 10mg po = 12.5mg IM CONVERTING FROM PO TO LONG-ACTING DECANOATE: Onset: 24-72hr (4-72hr) Long-Acting SC (12.5-100mg) (1-4 wks) Round to nearest 12.5mg Method 1: 1.25 X po daily dose = equiv decanoate dose; admin Q2-3wks. Cont ½ po daily dose X 1st few mths Cmax: 48-96hr 25mg/ml Method 2: ↑ decanoate dose over 4wks & ↓ po dose over 4-8wks as follows (accelerate taper for sx of EPS): t½: 6.8-9.6dy (single dose) ORAL DECANOATE (Administer Q 2 weeks) 15dy (14-100dy chronic administration) ORAL DOSE (mg/dy) ↓ DOSE OVER (wks) INITIAL DOSE (mg) TARGET DOSE (mg) DOSE OVER (wks) Steady State: 2mth (1.5-3mth) 5 4 6.25 6.25 0 Duration Action: 2wk (1-6wk) Elimination: Hepatic to inactive metabolites 10 4 6.25 12.5 4 Hemodialysis: Not dialyzable 20 8 6.25 12.5 4 30 8 6.25 25 4 40 8 6.25 25 4 Method 3: Admin equivalent decanoate dose Q2-3wks. -

Adverse Effects in Women: Implications for Drug Development and Regulatory Policies
Review For reprint orders, please contact [email protected] Adverse effects in women: implications for drug development and regulatory policies Expert Rev. Clin. Pharmacol. 4(4), 453–466 (2011) Ameeta Parekh†1, The requirement to establish safety of drugs prior to marketing has been in place since 1938 by Emmanuel O Fadiran1, the US Food, Drug and Cosmetic Act and is by no means a new concept. The efficacy regulations Kathleen Uhl2 were enacted in 1962 via the Kefauver–Harris Amendment and the drug approval process has and Douglas C evolved thereafter. The assessment of safety and efficacy of drug products is made by 2 pharmaceutical companies during drug development, which then goes through a regulatory Throckmorton review by the US FDA for the determination of market approval or nonapproval. The drug 1Food and Drug Administration, development and regulatory approval processes have endured close ongoing scrutiny by Office of Women’s Health, 10903 New Hampshire Avenue, regulatory bodies, the public, US Congress and academic and private organizations and, as a Silver Spring, MD 10993, USA result, have ensured continual refinement. Over the years, evidence has been emerging on varied 2Food and Drug Administration, drug responses in subgroup populations, and the underlying biology associated with age, race Center for Drug Evaluation and and sex as demographic variables have been examined. The resulting growing knowledge of Research, Office of Medical Policy, 10903 New Hampshire Avenue, disease burden, treatment response and disparate outcomes has generated opportunities to Silver Spring, MD 10993, USA streamline and improve treatment outcomes in these populations. This article discusses the †Author for correspondence: historical context of women’s participation in clinical drug trials submitted to the FDA for Tel.: +1 301 796 9445 regulatory review and approval purposes. -

Prediction of Premature Termination Codon Suppressing Compounds for Treatment of Duchenne Muscular Dystrophy Using Machine Learning
Prediction of Premature Termination Codon Suppressing Compounds for Treatment of Duchenne Muscular Dystrophy using Machine Learning Kate Wang et al. Supplemental Table S1. Drugs selected by Pharmacophore-based, ML-based and DL- based search in the FDA-approved drugs database Pharmacophore WEKA TF 1-Palmitoyl-2-oleoyl-sn-glycero-3- 5-O-phosphono-alpha-D- (phospho-rac-(1-glycerol)) ribofuranosyl diphosphate Acarbose Amikacin Acetylcarnitine Acetarsol Arbutamine Acetylcholine Adenosine Aldehydo-N-Acetyl-D- Benserazide Acyclovir Glucosamine Bisoprolol Adefovir dipivoxil Alendronic acid Brivudine Alfentanil Alginic acid Cefamandole Alitretinoin alpha-Arbutin Cefdinir Azithromycin Amikacin Cefixime Balsalazide Amiloride Cefonicid Bethanechol Arbutin Ceforanide Bicalutamide Ascorbic acid calcium salt Cefotetan Calcium glubionate Auranofin Ceftibuten Cangrelor Azacitidine Ceftolozane Capecitabine Benserazide Cerivastatin Carbamoylcholine Besifloxacin Chlortetracycline Carisoprodol beta-L-fructofuranose Cilastatin Chlorobutanol Bictegravir Citicoline Cidofovir Bismuth subgallate Cladribine Clodronic acid Bleomycin Clarithromycin Colistimethate Bortezomib Clindamycin Cyclandelate Bromotheophylline Clofarabine Dexpanthenol Calcium threonate Cromoglicic acid Edoxudine Capecitabine Demeclocycline Elbasvir Capreomycin Diaminopropanol tetraacetic acid Erdosteine Carbidopa Diazolidinylurea Ethchlorvynol Carbocisteine Dibekacin Ethinamate Carboplatin Dinoprostone Famotidine Cefotetan Dipyridamole Fidaxomicin Chlormerodrin Doripenem Flavin adenine dinucleotide -

Aspirin Intervention for the Reduction of Colorectal Cancer Risk (ASPIRED): a Study Protocol for a Randomized Controlled Trial David A
Drew et al. Trials (2017) 18:50 DOI 10.1186/s13063-016-1744-z STUDYPROTOCOL Open Access ASPirin Intervention for the REDuction of colorectal cancer risk (ASPIRED): a study protocol for a randomized controlled trial David A. Drew1,2, Samantha M. Chin1,2, Katherine K. Gilpin1,2, Melanie Parziale1,2, Emily Pond1,2, Madeline M. Schuck1,2, Kathleen Stewart2, Meaghan Flagg3, Crystal A. Rawlings3, Vadim Backman4, Peter J. Carolan2, Daniel C. Chung2, Francis P. Colizzo III2, Matthew Freedman5, Manish Gala1,2, John J. Garber2, Curtis Huttenhower6, Dmitriy Kedrin2, Hamed Khalili1,2, Douglas S. Kwon3, Sanford D. Markowitz8, Ginger L. Milne9, Norman S. Nishioka2, James M. Richter2, Hemant K. Roy10, Kyle Staller1,2, Molin Wang6,7,11 and Andrew T. Chan1,2,5,11,12* Abstract Background: Although aspirin is recommended for the prevention of colorectal cancer, the specific individuals for whom the benefits outweigh the risks are not clearly defined. Moreover, the precise mechanisms by which aspirin reduces the risk of cancer are unclear. We recently launched the ASPirin Intervention for the REDuction of colorectal cancer risk (ASPIRED) trial to address these uncertainties. Methods/design: ASPIRED is a prospective, double-blind, multidose, placebo-controlled, biomarker clinical trial of aspirin use in individuals previously diagnosed with colorectal adenoma. Individuals (n = 180) will be randomized in a 1:1:1 ratio to low-dose (81 mg/day) or standard-dose (325 mg/day) aspirin or placebo. At two study visits, participants will provide lifestyle, dietary and biometric data in addition to urine, saliva and blood specimens. Stool, grossly normal colorectal mucosal biopsies and cytology brushings will be collected during a flexible sigmoidoscopy without bowel preparation. -

Downloaded from Bioscientifica.Com at 09/25/2021 12:14:58PM Via Free Access
ID: 19-0149 8 6 Q Pang et al. PHO patients with soft tissue 8:6 736–744 giant tumor caused by HPGD mutation RESEARCH The first case of primary hypertrophic osteoarthropathy with soft tissue giant tumors caused by HPGD loss-of-function mutation Qianqian Pang1,2, Yuping Xu1,3, Xuan Qi1, Yan Jiang1, Ou Wang1, Mei Li1, Xiaoping Xing1, Ling Qin2 and Weibo Xia1 1Department of Endocrinology, Key Laboratory of Endocrinology, Ministry of Health, Peking Union Medical College Hospital, Chinese Academy of Medical Sciences, Beijing, China 2Musculoskeletal Research Laboratory and Bone Quality and Health Assessment Centre, Department of Orthopedics & Traumatology, The Chinese University of Hong Kong, Hong Kong SAR, Hong Kong 3Department of Endocrinology, The First Affiliated Hospital of Shanxi Medicalniversity, U Taiyuan, Shanxi, China Correspondence should be addressed to L Qin or W Xia: [email protected] or [email protected] Abstract Background: Primary hypertrophic osteoarthropathy (PHO) is a rare genetic multi-organic Key Words disease characterized by digital clubbing, periostosis and pachydermia. Two genes, f primary hypertrophic HPGD and SLCO2A1, which encodes 15-hydroxyprostaglandin dehydrogenase (15-PGDH) osteoarthropathy and prostaglandin transporter (PGT), respectively, have been reported to be related to f PHO PHO. Deficiency of aforementioned two genes leads to failure of prostaglandin E2 (PGE2) f HPGD mutation degradation and thereby elevated levels of PGE2. PGE2 plays an important role in f soft tumor tumorigenesis. Studies revealed a tumor suppressor activity of 15-PGDH in tumors, such f COX2 selective inhibitor treatment as lung, bladder and breast cancers. However, to date, no HPGD-mutated PHO patients presenting concomitant tumor has been documented. -

Neuroprotection by Chlorpromazine and Promethazine in Severe Transient and Permanent Ischemic Stroke
Mol Neurobiol (2017) 54:8140–8150 DOI 10.1007/s12035-016-0280-x Neuroprotection by Chlorpromazine and Promethazine in Severe Transient and Permanent Ischemic Stroke Xiaokun Geng1,2 & Fengwu Li1 & James Yip2 & Changya Peng2 & Omar Elmadhoun2 & Jiamei Shen1 & Xunming Ji1,3 & Yuchuan Ding1,2 Received: 29 June 2016 /Accepted: 31 October 2016 /Published online: 28 November 2016 # Springer Science+Business Media New York 2016 Abstract Previous studies have demonstrated depressive or enhance C + P-induced neuroprotection. C + P therapy im- hibernation-like roles of phenothiazine neuroleptics [com- proved brain metabolism as determined by increased ATP bined chlorpromazine and promethazine (C + P)] in brain levels and NADH activity, as well as decreased ROS produc- activity. This ischemic stroke study aimed to establish neuro- tion. These therapeutic effects were associated with alterations protection by reducing oxidative stress and improving brain in PKC-δ and Akt protein expression. C + P treatments con- metabolism with post-ischemic C + P administration. ferred neuroprotection in severe stroke models by suppressing Sprague-Dawley rats were subjected to transient (2 or 4 h) the damaging cascade of metabolic events, most likely inde- middle cerebral artery occlusion (MCAO) followed by 6 or pendent of drug-induced hypothermia. These findings further 24 h reperfusion, or permanent (28 h) MCAO without reper- prove the clinical potential for C + P treatment and may direct fusion. At 2 h after ischemia onset, rats received either an us closer towards the development of an efficacious neuropro- intraperitoneal (IP) injection of saline or two doses of C + P. tective therapy. Body temperatures, brain infarct volumes, and neurological deficits were examined.