Author Index
Total Page:16
File Type:pdf, Size:1020Kb

Load more
Recommended publications
-

Checklist of the Mammals of Indonesia
CHECKLIST OF THE MAMMALS OF INDONESIA Scientific, English, Indonesia Name and Distribution Area Table in Indonesia Including CITES, IUCN and Indonesian Category for Conservation i ii CHECKLIST OF THE MAMMALS OF INDONESIA Scientific, English, Indonesia Name and Distribution Area Table in Indonesia Including CITES, IUCN and Indonesian Category for Conservation By Ibnu Maryanto Maharadatunkamsi Anang Setiawan Achmadi Sigit Wiantoro Eko Sulistyadi Masaaki Yoneda Agustinus Suyanto Jito Sugardjito RESEARCH CENTER FOR BIOLOGY INDONESIAN INSTITUTE OF SCIENCES (LIPI) iii © 2019 RESEARCH CENTER FOR BIOLOGY, INDONESIAN INSTITUTE OF SCIENCES (LIPI) Cataloging in Publication Data. CHECKLIST OF THE MAMMALS OF INDONESIA: Scientific, English, Indonesia Name and Distribution Area Table in Indonesia Including CITES, IUCN and Indonesian Category for Conservation/ Ibnu Maryanto, Maharadatunkamsi, Anang Setiawan Achmadi, Sigit Wiantoro, Eko Sulistyadi, Masaaki Yoneda, Agustinus Suyanto, & Jito Sugardjito. ix+ 66 pp; 21 x 29,7 cm ISBN: 978-979-579-108-9 1. Checklist of mammals 2. Indonesia Cover Desain : Eko Harsono Photo : I. Maryanto Third Edition : December 2019 Published by: RESEARCH CENTER FOR BIOLOGY, INDONESIAN INSTITUTE OF SCIENCES (LIPI). Jl Raya Jakarta-Bogor, Km 46, Cibinong, Bogor, Jawa Barat 16911 Telp: 021-87907604/87907636; Fax: 021-87907612 Email: [email protected] . iv PREFACE TO THIRD EDITION This book is a third edition of checklist of the Mammals of Indonesia. The new edition provides remarkable information in several ways compare to the first and second editions, the remarks column contain the abbreviation of the specific island distributions, synonym and specific location. Thus, in this edition we are also corrected the distribution of some species including some new additional species in accordance with the discovery of new species in Indonesia. -

A Phylogeny and Timescale for Marsupial Evolution Based on Sequences for Five Nuclear Genes
J Mammal Evol DOI 10.1007/s10914-007-9062-6 ORIGINAL PAPER A Phylogeny and Timescale for Marsupial Evolution Based on Sequences for Five Nuclear Genes Robert W. Meredith & Michael Westerman & Judd A. Case & Mark S. Springer # Springer Science + Business Media, LLC 2007 Abstract Even though marsupials are taxonomically less diverse than placentals, they exhibit comparable morphological and ecological diversity. However, much of their fossil record is thought to be missing, particularly for the Australasian groups. The more than 330 living species of marsupials are grouped into three American (Didelphimorphia, Microbiotheria, and Paucituberculata) and four Australasian (Dasyuromorphia, Diprotodontia, Notoryctemorphia, and Peramelemorphia) orders. Interordinal relationships have been investigated using a wide range of methods that have often yielded contradictory results. Much of the controversy has focused on the placement of Dromiciops gliroides (Microbiotheria). Studies either support a sister-taxon relationship to a monophyletic Australasian clade or a nested position within the Australasian radiation. Familial relationships within the Diprotodontia have also proved difficult to resolve. Here, we examine higher-level marsupial relationships using a nuclear multigene molecular data set representing all living orders. Protein-coding portions of ApoB, BRCA1, IRBP, Rag1, and vWF were analyzed using maximum parsimony, maximum likelihood, and Bayesian methods. Two different Bayesian relaxed molecular clock methods were employed to construct a timescale for marsupial evolution and estimate the unrepresented basal branch length (UBBL). Maximum likelihood and Bayesian results suggest that the root of the marsupial tree is between Didelphimorphia and all other marsupials. All methods provide strong support for the monophyly of Australidelphia. Within Australidelphia, Dromiciops is the sister-taxon to a monophyletic Australasian clade. -

A Species-Level Phylogenetic Supertree of Marsupials
J. Zool., Lond. (2004) 264, 11–31 C 2004 The Zoological Society of London Printed in the United Kingdom DOI:10.1017/S0952836904005539 A species-level phylogenetic supertree of marsupials Marcel Cardillo1,2*, Olaf R. P. Bininda-Emonds3, Elizabeth Boakes1,2 and Andy Purvis1 1 Department of Biological Sciences, Imperial College London, Silwood Park, Ascot SL5 7PY, U.K. 2 Institute of Zoology, Zoological Society of London, Regent’s Park, London NW1 4RY, U.K. 3 Lehrstuhl fur¨ Tierzucht, Technical University of Munich, Alte Akademie 12, 85354 Freising-Weihenstephan, Germany (Accepted 26 January 2004) Abstract Comparative studies require information on phylogenetic relationships, but complete species-level phylogenetic trees of large clades are difficult to produce. One solution is to combine algorithmically many small trees into a single, larger supertree. Here we present a virtually complete, species-level phylogeny of the marsupials (Mammalia: Metatheria), built by combining 158 phylogenetic estimates published since 1980, using matrix representation with parsimony. The supertree is well resolved overall (73.7%), although resolution varies across the tree, indicating variation both in the amount of phylogenetic information available for different taxa, and the degree of conflict among phylogenetic estimates. In particular, the supertree shows poor resolution within the American marsupial taxa, reflecting a relative lack of systematic effort compared to the Australasian taxa. There are also important differences in supertrees based on source phylogenies published before 1995 and those published more recently. The supertree can be viewed as a meta-analysis of marsupial phylogenetic studies, and should be useful as a framework for phylogenetically explicit comparative studies of marsupial evolution and ecology. -

Aptian–Albian) of Texas and Oklahoma
Reappraisal of the tribosphenidan mammals from the Trinity Group (Aptian–Albian) of Texas and Oklahoma BRIAN M. DAVIS and RICHARD L. CIFELLI Davis, B.M. and Cifelli, R.L. 2011. Reappraisal of the tribosphenidan mammals from the Trinity Group (Aptian–Albian) of Texas and Oklahoma. Acta Palaeontologica Polonica 56 (3): 441–462. The Trinity therians have long been the focus of attempts to reconstruct the evolutionary history of higher mammals, es− pecially in the context of the development of tribospheny. In this paper, we update the taxonomy of the tribosphenidan taxa known from the Trinity Group and establish with more confidence the premolar/molar count in each. Many isolated specimens can be referred to a specific tooth locus. Additional diversity is revealed within the Deltatheroida, with the de− scription of an additional species of Oklatheridium; Pappotherium is here considered a likely metatherian based on the in− ferred presence of four molars, while Holoclemensia is a basal eutherian (the opposite of some traditional interpretations). The remainder of the genera, Kermackia and Slaughteria, cannot be allied with either of the living groups of tribo− sphenidan mammals using the available data. We identify strong morphological diversity within this assemblage of stem taxa, including modifications to the traditional tribosphenic occlusal pattern in Kermackia. Mammalian evolution at the base of the tribosphenidan radiation was complex, and this underscores the need for caution when interpreting the mor− phology and relationships of taxa known by incomplete material. Key words: Tribosphenida, Metatheria, Eutheria, Deltatheroida, Trinity Group, Early Cretaceous. Brian M. Davis [[email protected]] and Richard L. Cifelli [[email protected]], Department of Zoology and Sam Noble Oklahoma Museum of Natural History, University of Oklahoma, 2401 Chautauqua Ave, Norman, OK, 73072, USA. -

SEDIMENTATION of the BARUN GOYOT FORMATION (Plates XXXIV-XLII )
RYSZARD GRADZINSKI & TOMASZ JERZYKIEWICZ SEDIMENTATION OF THE BARUN GOYOT FORMATION (Plates XXXIV-XLII ) Contents C ontents Pa ge Introduction . 112 Geological setting 112 Stratigraphy . .. 114 Previous work .. .. ... .. 114 Redefinition of the lithostratigraphic divisions. 115 Barun Goyot Formation ... 116 Nemegt Formation. .. .. 116 Relation between the observed profiles . 117 Petrographic description . .. 118 Clay and silt-grade sediments 119 Sand-grade sediments . .. 119 Intraformational gravels . 124 Exotic gravels . 124 Principal sediment types . 125 Flat-bedded sandstone units. 125 Mega cross-stratified units . 127 Massive, "structureless" sandstones. 134 Diversely stratified sandstones . 134 Alternating claystones and sandstones 136 Sedimentological interpretation 136 Occurrence of organic remains 140 Depositional environment . 141 Conclusions 143 Appen dix . 143 References . 144 Abstract. - The Barun Goyot Formation (previously termed Lower Nemegt Beds) is composed of clastic continental sediments of red-beds type; it is probably of Campanian age. The thickness of the formation exceeds 110 m. It is overlain by the Nemegt Formation (previously termed Upper Nemegt Beds), probably of Maast richtian age; the passage between the two format ions is gradual. A formal redefinition of the two Iithostra tigraphic divisions is presented in the paper. Five principal sediment types are distinguished in the Barun Goyot Formation, displaying sedimentary features indicative of various conditions of sedimentation. The lower part of the exposed profile of the Barun Goyot Formation is characterized by mega cross-stratified units, interpreted as dune deposits; they are intertonguing with water-deposited sediments laid in interdune areas. Chan nel deposits, attributed to intermittent streams are subordinate; massive sandstones, probably of various origin are predominating. The upper part of the profile of the formation is characterized by the predominance of flat-bedded sandstone units which were probabl y deposited in an intermittently flooded takyr-like area. -

AMERICAN MUSEUM NOVITATES Published by Number 330 Thz Auerican Museum of NAITURAL HISTORY October 30, 1928 New York City
AMERICAN MUSEUM NOVITATES Published by Number 330 THz AuERicAN MusEUM oF NAITURAL HISTORY October 30, 1928 New York City 56.9,33 (117:51.7) AFFINITIES OF THE MONGOLIAN CRETACEOUS INSECTIVORES' BY GEORGE GAYLORD SIMPSON The unique series of Mesozoic mammal remains found by the Third Asiatic Expedition in Mongolia has now been completely de- scribed in a series of three papers.2 The affinities of the one known multituberculate, Djadochtatherium matthewi, were as thoroughly discussed as the material warrants in the first paper, and no additional remarks seem necessary. The relationships of the more important insectivores, however, were only briefly discussed in the second paper and a review of the evidence, especially including the important new details given in the third paper, suggests some modification and amplification of the views already presented. Not only are these mammal remains by far the most complete ever discovered in the Mesozoic, but they also occupy a very strategic position in time and in space which makes close scrutiny of their relationships essential. In time they occur in the Cretaceous, when, according to theories formed before their discovery and based largely on early Tertiary mammals, the differentiation of the placental orders should be in progress and not yet far advanced. In space they occur in Central Asia in or near the region which a number of students, especially Osborn and Matthew, have considered as an important center of radiation and probably the very one whence came the groups of mammals which appear to have entered North America and Europe suddenly at the beginning of the Tertiary and which must have been undergoing an important deployment during upper Cretaceous time. -

New Data on the Skull and Dentition in the Mongolian Late Cretaceous Eutherian Mammal Zalambdalestes
NEW DATA ON THE SKULL AND DENTITION IN THE MONGOLIAN LATE CRETACEOUS EUTHERIAN MAMMAL ZALAMBDALESTES JOHN R. WIBLE Section of Mammals, Carnegie Museum of Natural History, 5800 Baum Boulevard, Pittsburgh, PA 15206 e-mail: [email protected] MICHAEL J. NOVACEK Division of Paleontology, American Museum of Natural History e-mail: [email protected] GUILLERMO W. ROUGIER Department of Anatomical Sciences and Neurobiology, School of Medicine, University of Louisville, Louisville, KY 40292 e-mail: [email protected] BULLETIN OF THE AMERICAN MUSEUM OF NATURAL HISTORY CENTRAL PARK WEST AT 79TH STREET, NEW YORK, NY 10024 Number 281, 144 pp., 58 ®gures, 3 tables Issued January 9, 2004 Copyright q American Museum of Natural History 2004 ISSN 0003-0090 2 BULLETIN AMERICAN MUSEUM OF NATURAL HISTORY NO. 281 CONTENTS Abstract ....................................................................... 3 Introduction .................................................................... 3 Materials and Methods .......................................................... 5 History of Investigations ......................................................... 7 Comparative Morphology ....................................................... 17 Dentition ................................................................... 17 Upper Incisors ............................................................ 18 Upper Canine ............................................................. 21 Upper Premolars .......................................................... 22 Upper -

New Late Cretaceous Mammals of Southern Kazakhstan
New Late Cretaceous mammals of southern Kazakhstan ALEXANDER 0. AVERIANOV Averianov, A.O. 1997. New Late Cretaceous mammals of southern Kazakhstan. -Acta Palaeontologica Polonica 42,2,243-256. Mammalian remains from the lower part of the Darbasa Formation (lower Campanian) at the 'Grey Mesa' locality in the Alymtau Range, southern Kazakhstan, are described. They include ?Bulganbaatar sp. (Multituberculata), Deltatheridiurn nessovi, sp. n. @el- tatheroida), and four eutherians: an undeterminated ?otlestid kemalestoid (?Otlestidae), ?Alymlestes sp. (Zalambdalestidae), ?Aspanlestes sp. (Zhelestidae), and an undetermi- nated eutherian. This new Cretaceous fauna is most similar to that from the Djadokhta Formation in Mongolia and may tentatively confirm an early Campanian age for the latter. Key w o r d s : Mammalia, Cretaceous, Kazakhstan, Mongolia. Alexander 0.Averianov [[email protected]. ru], Zoological Institute, Russian Academy of Sciences, Universitetskaya nab. 1, 199034 Saint Petersburg, Russia. Introduction Mesozoic mammals from Kazakhstan are poorly known. The first Mesozoic mammal from Kazakhstan (and the first from the former USSR), found in 1962, is a mandibular fragment with heavily damaged molars, the holotype of Beleutinus orl~vi(?Zalamb- dalestidae), from the Santonian Bostobe Formation at Zhalmauz Well, Kyzyl-Orda Region (Bazhanov 1972). Since that time only three additional mammal remains have been recognized from the Cretaceous of Kazakhstan: a cervical vertebra of a relatively large, undeterminated mammal from the Zhalmauz locality (Nessov & Khisarova 1988); a dentary fragment with p5, ml-3, the holotype of Sorlestes kara (Zhelestidae), from the drilling core penetrating lower Turonian deposits near Ashchikol' Lake, Chimkent Region (Nessov 1993); and a lower molar (ml), the holotype of Alymlestes kielanae (Zalambdalestidae) from the lower part (Campanian) of the Darbasa Forma- tion at the 'Grey Mesa' locality in the Alymtau Range, Chimkent Region (Averianov & Nessov 1995). -

Palaeontologia Electronica CT Reconstructions and Relationships Of
Palaeontologia Electronica http://palaeo-electronica.org CT reconstructions and relationships of the Early Cretaceous tribosphenidan mammal, Slaughteria eruptens (Trinity Group Texas, USA) Dale A. Winkler, Louis L. Jacobs, Y. Kobayashi, and Michael J. Polcyn ABSTRACT Among the nine taxa of tribosphenidan (boreosphenidan) mammals named from the Early Cretaceous Trinity Group of Texas and Oklahoma, Slaughteria eruptens pro- vides unique information about the evolution of tooth replacement. High-resolution CT scans and SEM imagery elucidate the jaw anatomy of S. eruptens. Associated upper and lower dentitions from Asian Cretaceous mammals provide the basis for a statistical model to quantify the tooth size relationships of the often cited Trinity Group mammals Pappotherium pattersoni and Holoclemensia texana, which were named based upon upper molar teeth. Two groups of Trinity mammals are evident based on lower molar size. Most large teeth are referable to H. texana. The lower m1 of S. eruptens fits neatly into the size range predicted for lower molars of P. pattersoni, and it shares com- patible tooth cusp relationships. Slaughteria eruptens is regarded most parsimoniously as a junior synonym of P. pattersoni. Pappotherium pattersoni thus shares the alternate premolar replacement pattern of more primitive therian mammals and basal eutheri- ans, but lacks any hint of a submolariform last premolar, as typifies Eutheria. Other small therian mammal teeth from the Trinity Group should be evaluated as possible deciduous teeth. Dale A. Winkler. Department of Earth Sciences, Southern Methodist University, Dallas, TX 75275 USA. [email protected] Louis L. Jacobs. Department of Earth Sciences, Southern Methodist University, Dallas, TX 75275 USA. [email protected] Yoshi Kobayashi. -

Population of Northern Quoll Dasyurus Hallucatus
THE ECOLOGY OF AN INSULAR POPULATION OF NORTHERN QUOLL DASYURUS HALLUCATUS Jai Thomas BSc. Environmental Science (1st Class Hons) Submitted in fulfilment of the requirements for the degree of Doctor of Philosophy Environmental and Conservation Sciences Murdoch University 2020 Dasyurus hallucatus, Koolan Island. Illustration by E. Harradine. The ecology of an insular population of northern quoll Dasyurus hallucatus ii Abstract The northern quoll (Dasyurus hallucatus) is the smallest of the four quoll species endemic to Australia, and has suffered significant recent declines due to the spread of a number of threatening processes including ingestion of the toxic cane toad, altered fire regimes, and habitat loss and modification. In many areas, the species has survived by persisting in rocky country but populations are fragmented and isolated. In the recent mining boom of Western Australia many areas where the northern quoll has persisted were targeted for mining, and there have been an increasing number of mining projects needing to manage and mitigate impacts to populations of this species. There are also a number of conservation initiatives in progress that aim to buffer the northern quoll from the continuing threat of exotic species and habitat modification. However, the northern quoll is a cryptic species that occurs in remote areas and research to date has been limited by logistic constraints and sampling difficulty. On Koolan Island, the northern quoll has persisted despite a long history of mining for iron ore (1934 – 1940; 1965 - 1993; 2006 - present). Therefore, this thesis aimed to study the ecology of Koolan Island northern quolls to better understand the population dynamics, diet and habitat use of an island population, and examine any interactions with modified areas. -
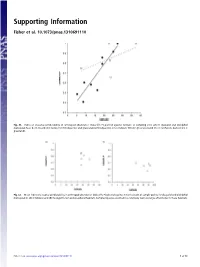
Supporting Information
Supporting Information Fisher et al. 10.1073/pnas.1310691110 Fig. S1. Index of seasonal predictability in arthropod abundance (Colwell’s P) plotted against latitude of sampling sites where dasyurid and didelphid marsupials have been recorded in rainforest (filled points) and grassland (unfilled points). Lines indicate fitted regressions (solid line = rainforest, dashed line = grassland). Fig. S2. Mean index of seasonal predictability in arthropod abundance (Colwell’s P) plotted against mean latitude of sample points for dasyurid and didelphid marsupials in (A) shrubland and (B) Eucalypt forest and woodland habitats. Sampled species occurred in a relatively narrow range of latitudes in these habitats. Fisher et al. www.pnas.org/cgi/content/short/1310691110 1of13 Fig. S3. Phylogeny of insectivorous marsupials with known life history data, based on ref. 1 with updates from ref. 2. 1. Cardillo M, Bininda-Emonds ORP, Boakes E, Purvis A (2004) A species-level phylogenetic supertree of marsupials. J Zool 264(1):11–31. 2. Fritz SA, Bininda-Emonds ORP, Purvis A (2009) Geographical variation in predictors of mammalian extinction risk: Big is bad, but only in the tropics. Ecol Lett 12(6):538–549. Fisher et al. www.pnas.org/cgi/content/short/1310691110 2of13 Fisher et al. Table S1. Reproductive traits and diets of insectivorous marsupials Latitude (south) www.pnas.org/cgi/content/short/1310691110 Proportion Proportion Female for Habitat of of age species class for females males Male Breeding Copulation Litters Scrotal at first with species Genus species -

The Mammals of Southern West Sepik Province, Papua New Guinea: Their Distribution, Abundance, Human Use and Zoogeography
AUSTRALIAN MUSEUM SCIENTIFIC PUBLICATIONS Flannery, Tim F., & Seri, L., 1990. The mammals of southern West Sepik Province, Papua New Guinea: their distribution, abundance, human use and zoogeography. Records of the Australian Museum 42(2): 173–208. [6 July 1990]. doi:10.3853/j.0067-1975.42.1990.114 ISSN 0067-1975 Published by the Australian Museum, Sydney naturenature cultureculture discover discover AustralianAustralian Museum Museum science science is is freely freely accessible accessible online online at at www.australianmuseum.net.au/publications/www.australianmuseum.net.au/publications/ 66 CollegeCollege Street,Street, SydneySydney NSWNSW 2010,2010, AustraliaAustralia Records of the Australian Museum (1990) Vol. 42: 173-208. ISSN 0067 1975 173 The Mammals of Southern West Sepik Province, Papua New Guinea: their Distribution, Abundance, Human Use and Zoogeography T.F. FLANNERyl & L. SERI2 IThe Australian Museum, 6 College Street, Sydney, NSW 2000, Australia 2Department of Environment and Conservation, Division of Wildlife, P.O. Box 6601, Boroko, Papua New Guinea ABSTRACT. A mammal survey was carried out between 1984 and 1987 in southern West Sepik Province, Papua New Guinea. Eleven major collecting localities, as well as some more minor ones, lying at altitudes of between 120 and 3,200 m were investigated. Voucher specimens for 87 indigenous mammal taxa were obtained, but research suggests that mammal diversity in the area may be as high as 120 species. This is the highest mammal diversity recorded anywhere in Australasia. A similar high bird diversity suggests that the area may be one of exceptionally high biodiversity overall. The most diverse mammal assemblages in the study area are found in the midmontane oak forests (between 1,500 and 2,500 m).