Melon Crops Improvement Through Biotechnological Techniques for the Changing Climatic Conditions of the 21 St Century
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

11 Available Online at 267
ISSN 2395-3411 Available online at www.ijpacr.com 267 _________________________________________________________Research Article Phytochemical Investigation and Evaluation of In-vitro Antilithiatic Activity of Muskmelon Fruit Syed Mumeena*, Ch. Srilakshmi, B. Tejeswini and T. Aruna Department of Pharmaceutical Management and Regulatory Affairs, Chalapathi Institute of Pharmaceutical Sciences, Lam, Guntur, Andhra Pradesh, India. __________________________________________________________________ ABSTRACT Muskmelon or Cucumis melo L. is one of the melon families that are widely consumed in many countries. The plant’s ability to adapt to many climate condition makes it available in the market throughout the year. Phytochemicals is one of the important components in plant. In this study, the methanolic extract of muskmelon fruit were studied and tested for antilithiatic activity. The fruit have ability to brake or dissolute the stones present in the urinary tract. Keywords: Muskmelon, urolithiasis, calcium oxalate, calcium phosphate, urinary stones. INTRODUCTION Urolithiasis is the word derived from Greek words Ouron (urine) and lithos (stone).Urolithiasis means formation of stone in the urinary system (calculi formed or located anywhere in the urinary system).1 It comprises Nephrolithiasis, formation of stones in Kidney; Ureterolithaisis, formation of stones in the ureter, and cystolithiasis formation of stones in bladder. Urinary tract stones 1.calcareous stones (calcium containing & radio-opaque) (75-90%) a) Calcium oxalate (whewellite & weddellite) b) Basic calcium phosphate 2. Non calcareous stones (non radio-opaque) 3. Struvite (Magnesium ammonium phosphate) (10-15%) 4. Uric acid (3-10%) 5. Cystine (0.5-1%) About 80% of those with kidney stones are men. Men most commonly experience their first episode between20-30years of age, while for women the age at first presentation is somewhat later. -

Correlating Sensory Attributes, Textural Parameters and Volatile Organic
CORRELATING SENSORY ATTRIBUTES, TEXTURAL PARAMETERS AND VOLATILE ORGANIC COMPOUNDS FOR THE ASSESSMENT OF DISTINCTIVE QUALITY TRAITS OF MELON AND PEACH FRUIT CULTIVARS Tiago Luís Cardoso Ferreira Pinhanços de Bianchi Per citar o enllaçar aquest document: Para citar o enlazar este documento: Use this url to cite or link to this publication: http://hdl.handle.net/10803/671744 ADVERTIMENT. L'accés als continguts d'aquesta tesi doctoral i la seva utilització ha de respectar els drets de la persona autora. Pot ser utilitzada per a consulta o estudi personal, així com en activitats o materials d'investigació i docència en els termes establerts a l'art. 32 del Text Refós de la Llei de Propietat Intel·lectual (RDL 1/1996). Per altres utilitzacions es requereix l'autorització prèvia i expressa de la persona autora. En qualsevol cas, en la utilització dels seus continguts caldrà indicar de forma clara el nom i cognoms de la persona autora i el títol de la tesi doctoral. No s'autoritza la seva reproducció o altres formes d'explotació efectuades amb finalitats de lucre ni la seva comunicació pública des d'un lloc aliè al servei TDX. Tampoc s'autoritza la presentació del seu contingut en una finestra o marc aliè a TDX (framing). Aquesta reserva de drets afecta tant als continguts de la tesi com als seus resums i índexs. ADVERTENCIA. El acceso a los contenidos de esta tesis doctoral y su utilización debe respetar los derechos de la persona autora. Puede ser utilizada para consulta o estudio personal, así como en actividades o materiales de investigación y docencia en los términos establecidos en el art. -

High Tunnel Melon and Watermelon Production
High Tunnel Melon and Watermelon Production University of Missouri Extension M173 Contents Author Botany 1 Lewis W. Jett, Division of Plant Sciences, University of Missouri-Columbia Cultivar selection 3 Editorial staff Transplant production 4 MU Extension and Agricultural Information Planting in the high tunnel 5 Dale Langford, editor Dennis Murphy, illustrator Row covers 6 On the World Wide Web Soil management and fertilization 6 Find this and other MU Extension publications on the Irrigation 7 Web at http://muextension.missouri.edu Pollination 7 Photographs Pruning 8 Except where noted, photographs are by Lewis W. Jett. Trellising 8 Harvest and yield 9 Marketing 10 Pest management 10 Useful references 14 Melon and watermelon seed sources 15 Sources of high tunnels (hoophouses) 16 For further information, address questions to College of Dr. Lewis W. Jett Agriculture Extension State Vegetable Crops Specialist Food and Natural Division of Plant Sciences Resources University of Missouri Columbia, MO 65211 Copyright 2006 by the University of Missouri Board of Curators E-mail: [email protected] College of Agriculture, Food and Natural Resources High Tunnel Melon and Watermelon Production igh tunnels are low-cost, passive, melo has several botanical subgroups (Table 1). solar greenhouses that use no fossil In the United States, reticulatus and inodorus are Hfuels for heating or venting (Figure commercially grown, while the remaining groups 1). High tunnels can provide many benefits to are grown for niche or local markets. horticulture crop producers: The cantaloupe fruit that most Americans • High tunnels are used to lengthen the are familiar with is not actually a true cantaloupe. -

Melon Top 100 September
Melon top 100 september click here to download 6 days ago The Billboard KPop HOT ranks the most popular songs in Korea based on streaming and sales data from leading music services in the. MelOn (Hangul: ; RR: Mellon) is a South Korean online music store (Most popular in Korea). Charts for Sep Melon Top By Fleur Inoue. Melon Top (Gayo) in September Unavailable songs on www.doorway.ru 92 songs. Play on. Sep Melon Top By Fleur Inoue. Melon Top (Gayo) in September Unavailable songs on www.doorway.ru 97 songs. Play on. Sep 23, Omg just went on Melon app bts is on top charts too for Melon ( 23 @pm) (yesterday. 10 4 Stream K-Pop Top (July 5, ), a playlist by Melon Sound from desktop or your mobile device. August MelOn Monthly Chart TOP Dance Category #1 Red Flavorpic. www.doorway.ru PM - 8 Sep Retweets; Likes. Billboard K-Town is an online Billboard column which reports on chart information for K-pop The list is exclusive of Korea K-Pop Hot data. Figures in red highlight indicate the highest rating received by K-pop artists on that chart. – Current week's. The Melon Music Awards is a major music awards show that is held annually in South Korea and organized by kakao M (a kakao company) through its online music store, Melon. Contents. 1 Host venues; 2 Judging criteria; 3 Main awards. Artist of the Year; Album of the Year; Song of the Year; Top 10 .. Retrieved September 20, Official Digital Thread (Gaon Index, AK-PAK, Melon Roofhits, Gaon [ RECORD] "Kiss and Make Up" becomes the highest song a K-Pop act has . -

Generate Evaluation Form
Great Lakes Fruit, Vegetable & Farm Market EXPO Michigan Greenhouse Growers EXPO December 5-7, 2017 DeVos Place Convention Center, Grand Rapids, MI Vine Crops Where: Grand Gallery (main level) Room E & F MI Recertification credits: 2 (1B, COMM CORE, PRIV CORE) OH Recertification credits: 1 (presentations as marked) CCA Credits: PM(1.0) CM(1.0) Moderator: Ben Phillips, Vegetable Extension Educator, MSU Extension, Saginaw, MI 9:00 am Integrated Management of Powdery Mildew and Gummy Stem Blight on Cucurbits: A Florida Perspective (OH: 2B, 0.5 hr) Gary Vallad, Plant Pathology Dept., Univ. of Florida 9:30 am Perspectives and Opportunities for Growing Orange Flesh and Specialty Melons Jonathan Schultheis, Dept. Horticultural Science, North Carolina State University 10:00 am Best Management Practices: Controlling Insects While Protecting Pollinators (OH: CORE, 0.5 hr) Rick Foster, Entomology Dept., Purdue Univ. 10:30 am Cover Cropping in Melons for Reduced Washing After Harvest Chris Gunter, Horticultural Science Dept., North Carolina State Univ. 11:00 am Session Ends Perspectives and Opportunities for Growing Orange Flesh and Specialty Melons Jonathan R. Schultheis North Carolina State University Department of Horticultural Science 2721 Founders Drive, 264 Kilgore Raleigh, NC 27695-7609 [email protected] Orange Flesh Melon Types: The landscape of orange flesh muskmelons (cantaloupes) has changed dramatically over the past five to ten years. Traditionally, there were two primary orange flesh melon types grown and sold; western types, which are fruits that range in size from about 3 to 5 pounds which are heavily netted and not sutured, and eastern types which have some netting, have a shorter shelf life but have a larger fruit size than a western melon, and tend to have a softer flesh with apparent more flavor than a western melon. -

Transformation of 'Galia' Melon to Improve Fruit
TRANSFORMATION OF ‘GALIA’ MELON TO IMPROVE FRUIT QUALITY By HECTOR GORDON NUÑEZ-PALENIUS A DISSERTATION PRESENTED TO THE GRADUATE SCHOOL OF THE UNIVERSITY OF FLORIDA IN PARTIAL FULFILLMENT OF THE REQUIREMENTS FOR THE DEGREE OF DOCTOR OF PHILOSOPHY UNIVERSITY OF FLORIDA 2005 Copyright 2005 by Hector Gordon Nuñez-Palenius This document is dedicated to the seven reasons in my life, who make me wake up early all mornings, work hard in order to achieve my objectives, dream on new horizons and goals, feel the beaty of the wind, rain and sunset, but mostly because they make me believe in God: my Dads Jose Nuñez Vargas and Salvador Federico Nuñez Palenius, my Moms Janette Ann Palenius Alberi and Consuelo Nuñez Solís, my wife Nélida Contreras Sánchez, my son Hector Manuel Nuñez Contreras and my daughter Consuelo Janette Nuñez Contreras. ACKNOWLEDGMENTS This dissertation could not have been completed without the support and help of many people who are gratefully acknowledged here. My greatest debt is to Dr. Daniel James Cantliffe, who has been a dedicated advisor and mentor, but mostly an excellent friend. He provided constant and efficient guidance to my academic work and research projects. This dissertation goal would not be possible without his insightful, invariable and constructive criticism. I thank Dr. Daniel J. Cantliffe for his exceptional course on Advanced Vegetable Production Techniques (HOS-5565) and the economic support for living expenses during my graduate education in UF. I extend my appreciation to my supervisory committee, Dr. Donald J. Huber, Dr. Harry J. Klee, and Dr. Donald Hopkins, for their academic guidance. -

Melons HOM-EN-KAS
Health and Learning Success Go Hand-in-Hand Farmers’ markets can help students learn how food travels from the farm to the plate. They also showcase diversity of fresh fruits and vegetables. Studies have Harvest shown that increasing students’ knowledge of fruits and vegetables may result in increased consumption. Use Harvest of the Month to teach students about farmers’ markets and show them how of the Health and Learningto lead aSuccess healthy, Go Hand-in-Hand active lifestyle. It links with core curricula and connects the classroom, cafeteria,Farmers’ home markets and community.can help students learn how food travels from the farm to the plate. They also showcase diversity of fresh fruits and vegetables. Studies have Harvest shown that increasing students’ knowledge of fruits and vegetables may result in increased consumption. Use Harvest of the Month to teach students Exploring Melonsabout farmers’ markets and show them how to lead a healthy, of Offeringth activitiese that allow students to experience melons using their senses active lifestyle. It links with core curricula and connects Month engages them in the learningthe classroom, process cafeteria, and creates home andincreased community. interest, awareness and Growing Healthy Students support for eating more fruits and vegetables. Tools: Exploring Melons n Three or more differentOffering varieties activities ofthat melons* allow students to experience melons using their senses Montn Knives, hcutting boardsengages and them serving in the plates learning (one process for each and group)creates increased interest, awareness and Growing Healthn Plasticy Students food service support gloves for (oneeating pair more per fruits student) and vegetables. n Small plates or bowlsTools: n Paper and pencilsn Three or more different varieties of melons* *Refer to Eat Your Colorsn onKnives, the next cutting page for boards varieties. -
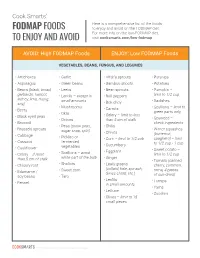
Fodmapfoods to Enjoy and Avoid
Cook Smarts’ Here is a comprehensive list of the foods FODMAP FOODS to enjoy and avoid on the FODMAP diet. For more info on the low-FODMAP diet, TO ENJOY AND AVOID visit cooksmarts.com/low-fodmap AVOID: High FODMAP Foods ENJOY: Low FODMAP Foods VEGETABLES, BEANS, FUNGUS, AND LEGUMES • Artichokes • Garlic • Alfalfa sprouts • Parsnips • Asparagus • Green beans • Bamboo shoots • Potatoes • Beans (black, broad, • Leeks • Bean sprouts • Pumpkin – garbanzo, haricot, • Lentils – except in • Bell peppers limit to 1/2 cup kidney, lima, mung, small amounts • Radishes soy) • Bok choy • Mushrooms • Scallions – limit to • Beets • Carrots • Okra green parts only • Black-eyed peas • Celery – limit to less • Onions than 5 cm of stalk • Seaweed – • Broccoli check ingredients • Peas (snow peas, • Chilis • Brussels sprouts • Winter squashes sugar snap, split) • Chives • Cabbage (butternut, • Pickled or • Corn – limit to 1/2 cob spaghetti) – limit • Cassava fermented • Cucumbers to 1/2 cup - 1 cup • Cauliflower vegetables • Eggplant • Sweet potato – • Celery – if more • Scallions – avoid limit to 1/2 cup white part of the bulb • Ginger than 5 cm of stalk • Tomato (canned, • Chicory root • Shallots • Leafy greens cherry, common, • Sweet corn (collard, kale, spinach, roma, 4 pieces • Edamame / Swiss chard, etc.) soy beans • Taro of sun-dried) • Lentils – • Turnips • Fennel in small amounts • Yams • Lettuce • Zucchini • Olives – limit to 15 small pieces © 2018 Cook Smarts | All Rights Reserved | Page 1 AVOID: High FODMAP Foods ENJOY: Low FODMAP Foods FRUITS -

Melon Match up Match the Melon Name and Description with Its Picture
MELON MADNESS! Vine growing fruit, melons are members of the Cucurbitaceous family, with their next closest relatives being squashes and cucumbers. Ancient Egyptians have been cultivating melons 2000 years BC, and were introduced to America by the Spanish settlers during the 15th and 16th century. There are numerous varieties of melons that differ in size, color and sweetness of their fruit. CLASSROOM DISCUSSION/ACTIVITY Melon Match Up Match the melon name and description with its picture. A B C D E 1. Gac Melon 2. 3. Horned Melon 4. Canary 5. Cantaloupe Gac is primarily a The most popular Watermelon It gets its name from Melon Southeast Asian melon in the United Containing 92% the horn-like spikes Named after the fruit. It is not very States, this melon water, the and has a texture that bright yellow abundant as it has a was first cultivated watermelon is is very similar to that of color of the short harvet season. in the 1700’s in the appropriately a cucumber. The taste canary bird, the The seeds of the fruit Italian village of names. The is tart-like, and often canary melon has are rich in flavor and Cantalup. largest recorded described as a a prominent are cooked with rice Cantaloupe is a rich watermelon combination of sweet flavor and in Vietnam. Its juice source for dietary weighed 262 cucumber and zucchini. tastes like is becoming popular fiber, vitamin A, pounds. cantaloupe. out side of Asia, as it vitamin C, iron, is very high in phosphorus and nutrients. 1. _____ 2.______ 3.______ 4.______ 5.______ potassium. -

Food Intolerance
TABLE OF CONTENTS FOOD INTOLERANCE ...............................................03 AESKULISA® Food Intolerance Line ...................04 ELISA TESTING Kit Components ..................................................07 OVERVIEW ...............................................................09 AESKULISA® Food Intolerance ONE G ..................09 ® AESKULISA Food Intolerance ONE G4 .................09 AESKULISA® Food Intolerance TWO G ..................09 ® AESKULISA Food Intolerance TWO G4 .................09 AESKULISA® Food Intolerance THREE G ...............09 ® AESKULISA Food Intolerance THREE G4 ..............09 AESKULISA® Food Intolerance FOUR G .................09 ® AESKULISA Food Intolerance FOUR G4 ................09 LISTS OF FOODS ......................................................10 PRODUCT HIGHLIGHTS .............................................15 AESKUBLOTS® Food Intolerance Line ............. 16 BLOT TESTING Kit Components ..................................................19 OVERVIEW ...............................................................21 AESKUBLOTS® Food Intolerance ONE ..................22 AESKUBLOTS® Food Intolerance TWO ..................22 AESKUBLOTS® Food Intolerance THREE ...............23 AESKUBLOTS® Food Intolerance FOUR .................23 AESKUBLOTS® Food Intolerance FIVE ..................24 AESKUBLOTS® Food Intolerance SIX ....................24 AESKUBLOTS® Food Intolerance SEVEN ...............24 LISTS OF FOODS ......................................................25 PRODUCT HIGHLIGHTS .............................................29 -

Alginate-Based Edible Coating to Enhance Quality And
ALGINATE-BASED EDIBLE COATING TO ENHANCE QUALITY AND EXTEND SHELF LIFE OF FRESH-CUT WATERMELON (CITRULLUS LANATUS) A Thesis by RABIA ESMA SIPAHI Submitted to the Office of Graduate Studies of Texas A&M University in partial fulfillment of the requirements for the degree of MASTER OF SCIENCE August 2012 Biological and Agricultural Engineering Alginate-based Edible Coating to Enhance Quality and Extend Shelf Life of Fresh- Cut Watermelon (Citrullus Lanatus) Copyright 2012 Rabia Esma Sipahi ALGINATE-BASED EDIBLE COATING TO ENHANCE QUALITY AND EXTEND SHELF LIFE OF FRESH-CUT WATERMELON (CITRULLUS LANATUS) A Thesis by RABIA ESMA SIPAHI Submitted to the Office of Graduate Studies of Texas A&M University in partial fulfillment of the requirements for the degree of MASTER OF SCIENCE Approved by: Chair of Committee, Elena Castell-Perez Committee Members, Rosana Moreira Carmen Gomes Alejandro Castillo Head of Department, Stephen W. Searcy August 2012 Biological and Agricultural Engineering iii ABSTRACT Alginate-based Edible Coating to Enhance Quality and Extend Shelf life of Fresh-Cut Watermelon (Citrullus Lanatus). (August 2012) Rabia Esma Sipahi, B.S., Ankara University Chair of Advisory Committee: Dr. Elena Castell-Perez Fresh-cut watermelon is appreciated for its taste, flavor, and juiciness. However, there are challenges in maintaining the freshness since fresh-cut processing of fruits promotes faster deterioration. Our objective was to determine the effectiveness of multilayered antimicrobial edible coating on the shelf-life of fresh-cut watermelon while keeping its original attributes for longer, without affecting its sensory properties. A set of solutions containing sodium alginate (0.5, 1, 2% w/w), beta- cyclodextrin, trans-cinnamaldehyde (antimicrobial agent), pectin, and calcium lactate were used as coating systems for fresh-cut watermelon cylinders. -
Melon & Watermelon
Melon & Watermelon Assortment 2019-2020 Meet our Melon & Watermelon team Dear customer, We are happy to present to you our new international melon booklet: an overview of our melon and watermelon varieties. Our product development activities in recent years have led us into new production areas and consumer markets. Hence, our range today includes varieties that are grown worldwide. Nevertheless, our international focus does not exclude new labels for local markets like Asian Jewels, a range of melon types and varieties specially bred for the various Asian markets. There are different ways of growing melons - e.g. open field or indoor, horizontal or vertical, local-for-local or further afield because of climate influences - but, ultimately, we all share the same goal: satisfied consumers − anywhere, anytime. Should you have any questions about this booklet, please feel free to contact your local Rijk Zwaan representative. Crop Team Melon & Watermelon Rijk Zwaan Crop coordinator Maarten den Hertog [email protected] Crop coordinator Diego Maestre Claramunt [email protected] Crop coordinator Vincent van Wolferen [email protected] 2 Americas Argentina Ignacio Damián Lopez [email protected] Brazil (melon) Weber Carlos Pinheiro [email protected] Brazil (watermelon) Eric Rodrigo Bezerra [email protected] Central America Darwin Lopez Acevedo [email protected] Chile Carlos Hurel Estay [email protected] Mexico Lionel Aragon Garcia [email protected] USA Joe Vandiver [email protected] USA