'Cripps Pink' Apples 2 1.3 Thesis Outline 3 1.4 References 5
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Applewood Focused on Apple Varieties
- Advertisement - Applewood focused on apple varieties August 21, 2019 Applewood Fresh Growers LLC is distinguishing itself with planning for the “next best managed varieties,” according to Nick Mascari, Applewood’s president. The Michigan grower-packer-shipper is testing apple varieties in the ground as part of the work toward serving the trade with the best of oncoming special new apple varieties. “We are really looking at making a statement with managed varieties and higher-end apples.” Mascari said that, for competitive reasons, “I can’t be specific” on what is being tested, but “we have five potential new varieties in test blocks.” If those are not ready for production now, “they will be further out. We are on the forefront for new sport varieties, and new strains of old varieties, atop having new managed varieties.” Antonia Mascari was recently promoted to become 1 / 3 the vice president of marketing for Applewood Fresh Growers LLC.With this, Applewood’s sales and marketing arm is “using more data to guide growers and customers” toward the best of the apples that will be coming from Michigan. By tracking data from all growing regions on apple sales for 52 weeks a year, growers can have the best information for planning the timing of new plantings and “we can help guide our customers to have the right assortment at the right times in their stores.” Mascari said USDA statistical data shows “We are the second largest grower-shipper in Michigan. Period. Our growth in volume is due to our team. We are pleased to be the leading sales company in Michigan.” “Our quality and volume will be up this year,” he continued. -

APPLE (Fruit Varieties)
E TG/14/9 ORIGINAL: English DATE: 2005-04-06 INTERNATIONAL UNION FOR THE PROTECTION OF NEW VARIETIES OF PLANTS GENEVA * APPLE (Fruit Varieties) UPOV Code: MALUS_DOM (Malus domestica Borkh.) GUIDELINES FOR THE CONDUCT OF TESTS FOR DISTINCTNESS, UNIFORMITY AND STABILITY Alternative Names:* Botanical name English French German Spanish Malus domestica Apple Pommier Apfel Manzano Borkh. The purpose of these guidelines (“Test Guidelines”) is to elaborate the principles contained in the General Introduction (document TG/1/3), and its associated TGP documents, into detailed practical guidance for the harmonized examination of distinctness, uniformity and stability (DUS) and, in particular, to identify appropriate characteristics for the examination of DUS and production of harmonized variety descriptions. ASSOCIATED DOCUMENTS These Test Guidelines should be read in conjunction with the General Introduction and its associated TGP documents. Other associated UPOV documents: TG/163/3 Apple Rootstocks TG/192/1 Ornamental Apple * These names were correct at the time of the introduction of these Test Guidelines but may be revised or updated. [Readers are advised to consult the UPOV Code, which can be found on the UPOV Website (www.upov.int), for the latest information.] i:\orgupov\shared\tg\applefru\tg 14 9 e.doc TG/14/9 Apple, 2005-04-06 - 2 - TABLE OF CONTENTS PAGE 1. SUBJECT OF THESE TEST GUIDELINES..................................................................................................3 2. MATERIAL REQUIRED ...............................................................................................................................3 -

Dusting Versus Spraying Apple Orchards in Ohio
BULLETIN 527 AUGUST, 1933 Dusting Versus Spraying Apple Orchards in Ohio F. H. Ballou and I. P. Lewis • OHIO AGRICULTURAL EXPERIMENT STATION Wooster, Ohio This page intentionally blank. DUSTING VERSUS SPRAYING APPLE ORCHARDS IN OHIO . INTRODUCTION F. :U:. BALLOU .AliD I. I'. LEWIS In response to a special request from the organized fruit growers of Ohio the Department of Horticulture of the Ohio Experiment Station began, in the spring of 1926, a series of experiments in dusting versus spraying apple orchards for prevention of injury to foliage and fruit by fungous diseases and insect pests. This series of dusting versus spraying experiments was carried on in eastern, central, northeastern, and southeastern Ohio. The five widely separated projects were located as follows: The Belmont County Experiment Farm, the Dale View Test Orchards (Licking County), the Mahoning County Experiment Farm, the Southeastern Experiment Farm (Meigs County), and the Washington County Experiment Farm. The orchards in Belmont, Licking, and Washington Counties are very favorably situated in hilly sections of the State and, for the most part, occupy areas of ground of such elevation as affords unusual frost protection to buds, blossoms, and newly set fruit. No losses of fruit from cold occurred in these orchards during the full term of the experiment. The orchard at the Mahoning County Experiment Farm occupies ground of moderate elevation, which affords fairly satisfactory cold-air drainage. In the spring of 1929 a partial loss of the apple crop was sustained by reason of freezing temperature immediately preceding the period of open blossoms. However, there was sufficient production of fruit to permit securing grades and counts in all of the variously dusted and sprayed plots. -
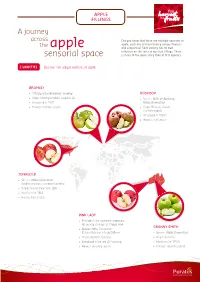
Discover Five Unique Varieties of Apple
APPLE FILLINGS A journey across Did you know that there are multiple varieties of apple, each one of them having unique flavours the apple and properties? Each variety has its own influence on the taste of our fruit fillings. There sensorial space is more to the apple story than at first appears. 5 VARIETIES Discover five unique varieties of apple: BRAMLEY • Officialy called Bramleys’ seeding BOSKOOP • Origin: Nottinghamshire, England, UK. • Species: Belle de Boskoop, • Introduced in 1837 Malus Domestica • Harvest: mid-late season • Origin: Boskoop, Gouda, The Netherlands • Introduced in 1850’s • Harvest: mid season JONAGOLD • Species: Malus Domestica (Golden delicious x Jonathan varieties) • Origin: Geneva, New York, USA • Introduced in 1943 • Harvest: late season PINK LADY • Pink lady is the (patented) trademark, the variety is known as Cripps Pink GRANNY SMITH • Species: Malus Domestica (Golden Delicious x Lady Williams) • Species: Malus Domestica • Origin: Western Australia • Origin: Australia • Introduced in the late 20th century • Introduced in 1860’s • Harvest: very-late season • Harvest: very-late season The flavour wheels show the flavour profiles of the fresh, unprocessed apples, that can easily FLAVOUR PROFILE be recognised in our different Topfils. SWEET sweet JONAGOLD red fruit notes The Jonagold is the sweetest variety cooked compote of this range. It has a very well- ripe fruit notes (pear, banana) balanced apple profile with some sour green and red fruit notes. green fruit notes (citrusy, unripe, fresh) astringent cider musty-woody PINK LADY The Pink Lady is slightly more sour than the Jonagold. It is very perfumy with specific pear/banana scents. These scents contribute to its nice ripe and juicy apple profile. -

Variety Description Origin Approximate Ripening Uses
Approximate Variety Description Origin Ripening Uses Yellow Transparent Tart, crisp Imported from Russia by USDA in 1870s Early July All-purpose Lodi Tart, somewhat firm New York, Early 1900s. Montgomery x Transparent. Early July Baking, sauce Pristine Sweet-tart PRI (Purdue Rutgers Illinois) release, 1994. Mid-late July All-purpose Dandee Red Sweet-tart, semi-tender New Ohio variety. An improved PaulaRed type. Early August Eating, cooking Redfree Mildly tart and crunchy PRI release, 1981. Early-mid August Eating Sansa Sweet, crunchy, juicy Japan, 1988. Akane x Gala. Mid August Eating Ginger Gold G. Delicious type, tangier G Delicious seedling found in Virginia, late 1960s. Mid August All-purpose Zestar! Sweet-tart, crunchy, juicy U Minn, 1999. State Fair x MN 1691. Mid August Eating, cooking St Edmund's Pippin Juicy, crisp, rich flavor From Bury St Edmunds, 1870. Mid August Eating, cider Chenango Strawberry Mildly tart, berry flavors 1850s, Chenango County, NY Mid August Eating, cooking Summer Rambo Juicy, tart, aromatic 16th century, Rambure, France. Mid-late August Eating, sauce Honeycrisp Sweet, very crunchy, juicy U Minn, 1991. Unknown parentage. Late Aug.-early Sept. Eating Burgundy Tart, crisp 1974, from NY state Late Aug.-early Sept. All-purpose Blondee Sweet, crunchy, juicy New Ohio apple. Related to Gala. Late Aug.-early Sept. Eating Gala Sweet, crisp New Zealand, 1934. Golden Delicious x Cox Orange. Late Aug.-early Sept. Eating Swiss Gourmet Sweet-tart, juicy Switzerland. Golden x Idared. Late Aug.-early Sept. All-purpose Golden Supreme Sweet, Golden Delcious type Idaho, 1960. Golden Delicious seedling Early September Eating, cooking Pink Pearl Sweet-tart, bright pink flesh California, 1944, developed from Surprise Early September All-purpose Autumn Crisp Juicy, slow to brown Golden Delicious x Monroe. -

Flesh Browning Disorder of 'Pink Lady' Apples
Th e Flesh Browning Disorder of ‘Pink Lady’™ Apples Hannah James1 and Jenny Jobling2 1Department of Horticulture, Cornell University, Ithaca, NY 2Applied Horticultural Research, Sydney, NSW, Australia he ‘Cripps Pink’ apple has become a popular vari- to be below the ety both in Australia where it was developed and acceptable lim- Tinternationally due to the distinctive fl avor charac- its for fi rmness teristics and at- and had also de- tractive appear- veloped an un- “Our research in Australia has shown that ance of the fruit. acceptable level Figure 1. Radial fl esh browning disorder (RFB) of the fl esh browning disorder of ‘Pink To establish a of skin greasi- ‘Pink Lady’™ apples in Australia. Lady’™ apples is three distinct disorders strong position ness prompting (radial fl esh browning, diff use fl esh among new and stricter quality guidelines to be enforced. In addition, a fl esh browning and CO2 injury.) Radial fl esh popular culti- browning disorder was detected; in 2003, 35 containers of browning was found to occur in warm vars, the ‘Cripps ‘Pink Lady’TM apples that had been exported from Australia growing districts and is characterized by Pink’ apple has to the United Kingdom were rejected due to the presence of browning of the vascular tissue of the been extensively the fl esh browning disorder, representing a large economic fruit, with the cortex tissue remaining marketed under loss to Australian apple growers and the potential loss of TM clear and relatively unaff ected. the trademarked the reputation of the ‘Pink Lady’ brand. TM Diff use fl esh browning was found to name ‘Pink La- Th e fl esh browning disorder of ‘Pink Lady’ apples TM occur in cool growing districts and is dy ’. -

Comprehensive Assessment of Organic Apple Production in Vermont: Experience from Two Orchard Systems, 2006-2013 Terence L
University of Vermont ScholarWorks @ UVM Graduate College Dissertations and Theses Dissertations and Theses 2015 Comprehensive Assessment of Organic Apple Production in Vermont: Experience from Two Orchard Systems, 2006-2013 Terence L. Bradshaw University of Vermont Follow this and additional works at: https://scholarworks.uvm.edu/graddis Part of the Agricultural Economics Commons, and the Horticulture Commons Recommended Citation Bradshaw, Terence L., "Comprehensive Assessment of Organic Apple Production in Vermont: Experience from Two Orchard Systems, 2006-2013" (2015). Graduate College Dissertations and Theses. 327. https://scholarworks.uvm.edu/graddis/327 This Dissertation is brought to you for free and open access by the Dissertations and Theses at ScholarWorks @ UVM. It has been accepted for inclusion in Graduate College Dissertations and Theses by an authorized administrator of ScholarWorks @ UVM. For more information, please contact [email protected]. COMPREHENSIVE ASSESSMENT OF ORGANIC APPLE PRODUCTION IN VERMONT: EXPERIENCE FROM TWO ORCHARD SYSTEMS, 2006-2013 A Dissertation Presented by Terence L. Bradshaw to The Faculty of the Graduate College of The University of Vermont In Partial Fulfillment of the Requirements for the Degree of Doctor of Philosophy Specializing in Plant and Soil Science May, 2015 Defense Date: March 25, 2014 Dissertation Examination Committee: Josef H. Gorres, Ph. D., Advisor Robert L. Parsons, Ph.D., Chairperson Lorraine P. Berkett, Ph.D. Sidney C. Bosworth, Ph.D. Cynthia J. Forehand, Ph.D., Dean of the Graduate College ABSTRACT Despite substantial consumer demand and willingness to pay premium prices for organically grown fruit, apple growers in Vermont and other New England states have been slow to adopt certified organic practices. -

Germplasm Sets and Standardized Phenotyping Protocols for Fruit Quality Traits in Rosbreed
Germplasm Sets and Standardized Phenotyping Protocols for Fruit Quality Traits in RosBREED Jim Luby, Breeding Team Leader Outline of Presentation RosBREED Demonstration Breeding Programs Standardized Phenotyping Protocols Reference Germplasm Sets SNP Detection Panels Crop Reference Set Breeding Pedigree Set RosBREED Demonstration Breeding Programs Clemson U WSU Texas A&M UC Davis U Minn U Arkansas Rosaceae Cornell U WSU MSU MSU Phenotyping Affiliates USDA-ARS Driscolls Corvallis Univ of Florida UNH Standardized Phenotyping Protocols Traits and Standardized Phenotyping Protocols • Identify critical fruit quality traits and other important traits • Develop standardized phenotyping protocols to enable data pooling across locations/institutions • Protocols available at www.RosBREED.org Apple Standardized Phenotyping Firmness, Crispness – Instrumental, Sensory Sweetness, Acidity – Intstrumental, Sensory Color, Appearance, Juiciness, Aroma – Sensory At harvest Cracking, Russet, Sunburn Storage 10w+7d Storage 20w+7d Maturity Fruit size 5 fruit (reps) per evaluation Postharvest disorders Harvest date, Crop, Dropping RosBREED Apple Phenotyping Locations Wenatchee, WA St Paul, MN Geneva, NY • One location for all evaluations would reduce variation among instruments and evaluators • Local evaluations more sustainable and relevant for future efforts at each institution • Conduct standardized phenotyping of Germplasm Sets at respective sites over multiple (2-3) seasons • Collate data in PBA format, conduct quality control, archive Reference -

National Apple Orchard Census 2012
National Apple Orchard Census 2012 1 Table of Contents Introduction Page 3 Executive Summary Page 4 Section 1 Orchard Area, Sales, Growers and Markets Page 6 Section 2 Varieties and Orchard Age Page 15 Section 3 Employment, Facilities, Business Development and Producer Page 21 Profile Appendix Tables Table 1: Apple Production Area (hectares) by Year of Planting and Apple Type in Ireland in 2012 Table 2: Number of Apple Trees by Year of Planting and Apple Type in Ireland in 2012 Table 3: Number of Growers of Each Apple Type by County in Ireland in 2012 Table 4: Sales of 2011 Harvest by Market by County Table 5: Production Area, Number of Growers and Sales for Most Commonly Grown Apple Varieties in Ireland in 2011/12 Table 6: Average (median) Prices per Tonne by Market for 2011 harvest Table 7: Apple Production Area (hectares) in Ireland in 1980-2012 2 National Apple Orchard Census 2012 Introduction The National Apple Orchard Census 2012 was compiled from information provided by the 45 commercial apple growers operating in the Republic of Ireland in 2012. The census provides an important overview of apple production, and comparisons are made to previous censuses (2007, 2002 and 1997) where possible. The census surveyed all commercial orchards, defined as follows: - a production area of at least 0.2 hectares of apples, including cider apples - the fruit from the orchard is sold every year - the orchard is sprayed against pests/diseases every year, or maintained to organic standards Within the 45 growers, there are two organic growers. Production areas are based on areas planted up to March 2012. -

The Pathogenicity and Seasonal Development of Gymnosporangium
Iowa State University Capstones, Theses and Retrospective Theses and Dissertations Dissertations 1931 The ap thogenicity and seasonal development of Gymnosporangium in Iowa Donald E. Bliss Iowa State College Follow this and additional works at: https://lib.dr.iastate.edu/rtd Part of the Agriculture Commons, Botany Commons, and the Plant Pathology Commons Recommended Citation Bliss, Donald E., "The ap thogenicity and seasonal development of Gymnosporangium in Iowa " (1931). Retrospective Theses and Dissertations. 14209. https://lib.dr.iastate.edu/rtd/14209 This Dissertation is brought to you for free and open access by the Iowa State University Capstones, Theses and Dissertations at Iowa State University Digital Repository. It has been accepted for inclusion in Retrospective Theses and Dissertations by an authorized administrator of Iowa State University Digital Repository. For more information, please contact [email protected]. INFORMATION TO USERS This manuscript has been reproduced from the microfilm master. UMl films the text directly from the original or copy submitted. Thus, some thesis and dissertation copies are in typewriter face, while others may be from any type of computer printer. The quality of this reproduction is dependent upon the quality of the copy submitted. Broken or indistinct print, colored or poor quality illustrations and photographs, print bleedthrough, substandard margins, and improper alignment can adversely affect reproduction. In the unlikely event that the author did not send UMl a complete manuscript and there are missing pages, these will be noted. Also, if unauthorized copyright material had to be removed, a note will indicate the deletion. Oversize materials (e.g.. maps, drawings, charts) are reproduced by sectioning the original, beginning at the upper left-hand comer and continuing from left to right in equal sections with small overiaps. -

Survey of Apple Clones in the United States
Historic, archived document Do not assume content reflects current scientific knowledge, policies, or practices. 5 ARS 34-37-1 May 1963 A Survey of Apple Clones in the United States u. S. DFPT. OF AGRffini r U>2 4 L964 Agricultural Research Service U.S. DEPARTMENT OF AGRICULTURE PREFACE This publication reports on surveys of the deciduous fruit and nut clones being maintained at the Federal and State experiment stations in the United States. It will b- published in three c parts: I. Apples, II. Stone Fruit. , UI, Pears, Nuts, and Other Fruits. This survey was conducted at the request of the National Coor- dinating Committee on New Crops. Its purpose is to obtain an indication of the volume of material that would be involved in establishing clonal germ plasm repositories for the use of fruit breeders throughout the country. ACKNOWLEDGMENT Gratitude is expressed for the assistance of H. F. Winters of the New Crops Research Branch, Crops Research Division, Agricultural Research Service, under whose direction the questionnaire was designed and initial distribution made. The author also acknowledges the work of D. D. Dolan, W. R. Langford, W. H. Skrdla, and L. A. Mullen, coordinators of the New Crops Regional Cooperative Program, through whom the data used in this survey were obtained from the State experiment stations. Finally, it is recognized that much extracurricular work was expended by the various experiment stations in completing the questionnaires. : CONTENTS Introduction 1 Germany 298 Key to reporting stations. „ . 4 Soviet Union . 302 Abbreviations used in descriptions .... 6 Sweden . 303 Sports United States selections 304 Baldwin. -

Handling of Apple Transport Techniques and Efficiency Vibration, Damage and Bruising Texture, Firmness and Quality
Centre of Excellence AGROPHYSICS for Applied Physics in Sustainable Agriculture Handling of Apple transport techniques and efficiency vibration, damage and bruising texture, firmness and quality Bohdan Dobrzañski, jr. Jacek Rabcewicz Rafa³ Rybczyñski B. Dobrzañski Institute of Agrophysics Polish Academy of Sciences Centre of Excellence AGROPHYSICS for Applied Physics in Sustainable Agriculture Handling of Apple transport techniques and efficiency vibration, damage and bruising texture, firmness and quality Bohdan Dobrzañski, jr. Jacek Rabcewicz Rafa³ Rybczyñski B. Dobrzañski Institute of Agrophysics Polish Academy of Sciences PUBLISHED BY: B. DOBRZAŃSKI INSTITUTE OF AGROPHYSICS OF POLISH ACADEMY OF SCIENCES ACTIVITIES OF WP9 IN THE CENTRE OF EXCELLENCE AGROPHYSICS CONTRACT NO: QLAM-2001-00428 CENTRE OF EXCELLENCE FOR APPLIED PHYSICS IN SUSTAINABLE AGRICULTURE WITH THE th ACRONYM AGROPHYSICS IS FOUNDED UNDER 5 EU FRAMEWORK FOR RESEARCH, TECHNOLOGICAL DEVELOPMENT AND DEMONSTRATION ACTIVITIES GENERAL SUPERVISOR OF THE CENTRE: PROF. DR. RYSZARD T. WALCZAK, MEMBER OF POLISH ACADEMY OF SCIENCES PROJECT COORDINATOR: DR. ENG. ANDRZEJ STĘPNIEWSKI WP9: PHYSICAL METHODS OF EVALUATION OF FRUIT AND VEGETABLE QUALITY LEADER OF WP9: PROF. DR. ENG. BOHDAN DOBRZAŃSKI, JR. REVIEWED BY PROF. DR. ENG. JÓZEF KOWALCZUK TRANSLATED (EXCEPT CHAPTERS: 1, 2, 6-9) BY M.SC. TOMASZ BYLICA THE RESULTS OF STUDY PRESENTED IN THE MONOGRAPH ARE SUPPORTED BY: THE STATE COMMITTEE FOR SCIENTIFIC RESEARCH UNDER GRANT NO. 5 P06F 012 19 AND ORDERED PROJECT NO. PBZ-51-02 RESEARCH INSTITUTE OF POMOLOGY AND FLORICULTURE B. DOBRZAŃSKI INSTITUTE OF AGROPHYSICS OF POLISH ACADEMY OF SCIENCES ©Copyright by BOHDAN DOBRZAŃSKI INSTITUTE OF AGROPHYSICS OF POLISH ACADEMY OF SCIENCES LUBLIN 2006 ISBN 83-89969-55-6 ST 1 EDITION - ISBN 83-89969-55-6 (IN ENGLISH) 180 COPIES, PRINTED SHEETS (16.8) PRINTED ON ACID-FREE PAPER IN POLAND BY: ALF-GRAF, UL.