Structural Insights Into a Plasmodium Falciparum IMC1 Protein Using
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Plasmodium Falciparum Is Not As Lonely As Previously Considered
AUTOPHAGIC PUNCTUM ARTICLE ADDENDUM Virulence 2:1, 71-76; January/February 2011; © 2011 Landes Bioscience Plasmodium falciparum is not as lonely as previously considered Franck Prugnolle,1,* Francisco Ayala,2 Benjamin Ollomo,3 Céline Arnathau,1 Patrick Durand1 and François Renaud1,* 1Laboratoire MIVEGEC; UM1-CNRS 5290-IRD 224, IRD Montpellier, France; 2Department of Ecology and Evolutionary Biology; University of California; Irvine, CA USA; 3Centre International de Recherches Médicales de Franceville; Franceville, Gabon ntil very recently, only one species The identification of Plasmodium spe- U(P. reichenowi) was known to be a cies circulating in great apes in Africa phylogenetic sister lineage of P. falciparum, was primarily done during the first half the main malignant agent of human of the twentieth century, on the basis of malaria. In 2009 and 2010, new studies morphological features.1 This approach have revealed the existence of several new has several limitations.4 First, phenotypic phylogenetic species related to this deadly plasticity can lead to incorrect identifica- parasite and infecting chimpanzees and tions. Second, morphological keys are gorillas in Africa. These discoveries invite often effective only for a particular life us to explore a whole set of new questions, stage which cannot always be observed which we briefly do in this article. or is difficult to be. Finally, and perhaps most important, this approach overlooks The Plasmodium species infecting morphologically cryptic taxa. These limi- humans and non-human primates cluster tations, together with the difficulty to into two distinct phylogenetic lineages collect and manipulate great apes, were (Fig. 1). One of these lineages (in yellow certainly, at least in part, responsible for in Fig. -

Legionella Genus Genome Provide Multiple, Independent Combinations for Replication in Human Cells
Supplemental Material More than 18,000 effectors in the Legionella genus genome provide multiple, independent combinations for replication in human cells Laura Gomez-Valero1,2, Christophe Rusniok1,2, Danielle Carson3, Sonia Mondino1,2, Ana Elena Pérez-Cobas1,2, Monica Rolando1,2, Shivani Pasricha4, Sandra Reuter5+, Jasmin Demirtas1,2, Johannes Crumbach1,2, Stephane Descorps-Declere6, Elizabeth L. Hartland4,7,8, Sophie Jarraud9, Gordon Dougan5, Gunnar N. Schroeder3,10, Gad Frankel3, and Carmen Buchrieser1,2,* Table S1: Legionella strains analyzed in the present study Table S2: Type IV secretion systems predicted in the genomes analyzed Table S3: Eukaryotic like domains identified in the Legionella proteins analyzed Table S4: Small GTPases domains detected in the genus Legionella as defined in the CDD ncbi domain database Table S5: Eukaryotic like proteins detected in the Legionella genomes analyzed in this study Table S6: Aminoacid identity of the Dot/Icm components in Legionella species with respect to orthologous proteins in L. pneumophila Paris Table S7: Distribution of seventeen highly conserved Dot/Icm secreted substrates Table S8: Comparison of the effector reperotoire among strains of the same Legionella species Table S9. Number of Dot/Icm secreted proteins predicted in each strain analyzed Table S10: Replication capacity of the different Legionella species analyzed in this study and collection of literature data on Legionella replication Table S11: Orthologous table for all genes of the 80 analyzed strains based on PanOCT. The orthologoss where defined with the program PanOCT using the parameters previously indicated in material and methods.) Figure S1: Distribution of the genes predicted to encode for the biosynthesis of flagella among all Legionella species. -
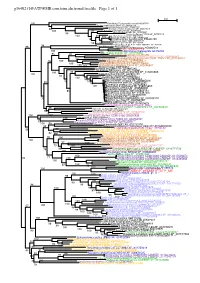
Page 1 G16482.T1@[email protected]
g16482.t1@[email protected] Page 1 of 1 0.2 100 Candidatus Thiomargarita nelsonii KHD07353 Bacillus thuringiensis Bt407 YP_006926419 Legionella pneumophila WP_014842957 88 Endozoicomonas montiporae WP_034879478 Acinetobacter baumannii WP_031968362 100 87 Vibrio nigripulchritudo WP_004402447 98 Psychrobacter sp. P11G3 WP_057760119 100 99 Xenorhabdus bovienii WP_038203911 100 Rouxiella chamberiensis WP_045046329 18 Dendroctonus ponderosae ERL83390 Serratia sp. M24T3 WP_009636677 98 56 100Lonsdalea quercina WP_026743324 Escherichia coli str. K-12 substr. MG1655 NP_415996 86 98 Chlorobi bacterium OLB5 KXK50071 CoptotermesTric_AAA92714 formosanus AGM32510 100 Histomonas meleagridis ACI16482 100Eucalyptus grandis XP_010033838 95 98 Ectocarpus siliculosus CBJ25596 Guillardia theta CCMP2712 XP_005838347 Phaeodactylum tricornutum CCAP 1055/1 XP_002180331 99 Aphanomyces astaci XP_009825348 95 100 Albugo laibachii Nc14 CCA15917 100 Phytophthora parasitica P1569 ETI54450 100 Phytophthora infestans T30-4 XP_002896607 100 Brugia malayi CDP93853 Bombus impatiens XP_003487345 Trichoplax adhaerens XP_002116081 95 Apteryx australis mantelli XP_013810665 100 100 Callorhinchus milii XP_007891153 Homo sapiens AAC50613 93 Callorhinchus milii XP_007900551 100 Clupea harengus XP_012674444 99 Chrysochloris asiatica XP_006865016 82 76 100 84Nannospalax galili XP_008842357 98 Cricetulus griseus XP_007626683 100 65Rousettus aegyptiacus XP_016015809 Condylura cristata XP_004683330 58Chinchilla lanigera XP_013367523 85 61Microcebus murinus XP_012594174 -

Inferring Natural Selection Signals in Plasmodium Vivax-Encoded Proteins Having a Potential Role in Merozoite Invasion
Identificación de señales de selección natural en genes de Plasmodium vivax que codifican proteínas involucradas en el proceso de invasión para determinar su potencial uso en una vacuna antimalárica. Diego Edison Garzón Ospina Tesis Doctoral presentada como requisito para optar al título de Doctor en Ciencias Biomédicas y Biológicas de la Universidad del Rosario Bogotá, 2018 1 Identificación de señales de selección natural en genes de Plasmodium vivax que codifican proteínas involucradas en el proceso de invasión para determinar su potencial uso en una vacuna antimalárica. Estudiante Diego Edison Garzón Ospina Biólogo, Universidad INCCA de Colombia Magister en Ciencias-Microbiología, Universidad Nacional de Colombia Director Manuel Alfonso Patarroyo Gutiérrez M.D., Dr.Sc. Jefe del Departamento de Biología Molecular e Inmunología Fundación Instituto de Inmunología de Colombia (FIDIC) Profesor Titular, Escuela de Medicina y Ciencias de la Salud Universidad del Rosario DOCTORADO EN CIENCIAS BIOMÉDICAS Y BIOLÓGICAS UNIVERSIDAD DEL ROSARIO Bogotá, 2018 2 AGRADECIMIENTOS Quiero dar mis agradecimientos y dedicar este trabajo a mi madre: Martha Ospina Vargas, y a mis hermanos: Yamile Garzón Ospina y Julián David Escobar Ospina, quienes me han acompañado y apoyado en todo momento. A Sindy Paola Buitrago Puentes por su acompañamiento y apoyo durante este tiempo. También agradecer al Dr. Manuel Alfonso Patarroyo Gutiérrez, por haberme acogido en su equipo de trabajo y por apoyarme a lo largo de todos estos años. Quiero agradecer y reconocer la labor de: Andrea Estefanía Ramos, Darwin Andrés Moreno Pérez, Elizabeth Gutiérrez Vásquez, Heidy Daniela Ortiz Suarez, Lady Johanna Forero Rodríguez, Laura Alejandra Ricaurte Contreras, Leidy Paola Reyes, Luis Alfredo Baquero, Paola Andrea Camargo Ayala, Sindy Paola Buitrago Puentes, Ricardo De León Montero y Yimara Grosso Paz, quienes aportaron su tiempo, dedicación y esfuerzo, permitiendo la culminación de este trabajo. -

Debugging Parasite Genomes: Using Metabolic Modeling to Accelerate Antiparasitic Drug Development
Debugging parasite genomes: Using metabolic modeling to accelerate antiparasitic drug development Maureen A. Carey Charlottesville, Virginia Bachelors of Science, Lafayette College 2014 A Dissertation presented to the Graduate Faculty of the University of Virginia in Candidacy for the Degree of Doctor of Philosophy Department of Microbiology, Immunology, and Cancer Biology University of Virginia September, 2018 i M. A. Carey ii Abstract: Eukaryotic parasites, like the casual agent of malaria, kill over one million people around the world annually. Developing novel antiparasitic drugs is a pressing need because there are few available therapeutics and the parasites have developed drug resistance. However, novel drug targets are challenging to identify due to poor genome annotation and experimental challenges associated with growing these parasites. Here, we focus on computational and experimental approaches that generate high-confidence hypotheses to accelerate labor-intensive experimental work and leverage existing experimental data to generate new drug targets. We generate genome-scale metabolic models for over 100 species to develop a parasite knowledgebase and apply these models to contextualize experimental data and to generate candidate drug targets. M. A. Carey iii Figure 0.1: Image from blog.wellcome.ac.uk/2010/06/15/of-parasitology-and-comics/. Preamble: Eukaryotic single-celled parasites cause diseases, such as malaria, African sleeping sickness, diarrheal disease, and leishmaniasis, with diverse clinical presenta- tions and large global impacts. These infections result in over one million preventable deaths annually and contribute to a significant reduction in disability-adjusted life years. This global health burden makes parasitic diseases a top priority of many economic development and health advocacy groups. -

Place De La Biologie Moléculaire Dans L'épidémiologie, Le Diagnostic Et L'évaluation De La Chimiorésistance Du Paludi
Place de la biologie moléculaire dans l’épidémiologie, le diagnostic et l’évaluation de la chimiorésistance du paludisme en République Démocratique du Congo Dieudonné Mvumbi Makaba, MD, MSc. Département des Sciences de Base Faculté de Médecine Université de Kinshasa Thèse soutenue et défendue publiquement en vue de l'obtention du grade de Docteur en Sciences Biomédicales (PhD) Promoteur: Co-promoteur: Marie-Pierre Hayette, PhD Jean-Marie Kayembe, PhD Ulg Unikin Thèse Place de la biologie moléculaire dans l’épidémiologie, le diagnostic et l’évaluation de la chimiorésistance du paludisme en République Démocratique du Congo Par Dieudonné Mvumbi Makaba Présentée et soutenue publiquement en vue de l'obtention du grade de Docteur en Sciences Biomédicales (PhD) Le 11 février 2017 Composition du Jury : 1. Professeur Marie-Pierre Hayette, Université de Liège (promoteur) 2. Professeur Jean-Marie Kayembe, Université de Kinshasa (co-promoteur) 3. Professeur Hippolyte Situakibanza, Université de Kinshasa 4. Professeur Gauthier Mesia, Université de Kinshasa 5. Professeur Patrick De Mol, Université de Liège 6. Professeur Dieudonné Mumba, Université de Kinshasa 7. Professeur Prosper Lukusa, Katholieke Universiteit Leuven Université de Kinshasa Faculté de Médecine Département des Sciences de Base Service de Biologie Moléculaire Place de la biologie moléculaire dans l’épidémiologie, le diagnostic et l’évaluation de la chimiorésistance du paludisme en République Démocratique du Congo Dieudonné Mvumbi Makaba Promoteur Co-promoteur Marie-Pierre Hayette, PhD Jean-Marie Kayembe N., PhD A ma famille … Table des matières Liste des figures iii Liste des tableaux iv Liste des abréviations v Remerciements vi Résumé ix Introduction………………………………………………………….1 Partie I: Etat des connaissances……………………………………3 I.1. -

Ovale Wallikeri
A New Real-Time PCR for the Detection of Plasmodium ovale wallikeri Adriana Calderaro1*, Giovanna Piccolo1, Chiara Gorrini1, Sara Montecchini1, Sabina Rossi1, Maria Cristina Medici1, Carlo Chezzi1, Georges Snounou2,3 1 Department of Pathology and Laboratory Medicine, Section of Microbiology, University of Parma, Parma, Italy, 2 Universite´ Pierre et Marie Curie - Paris VI, UMR S 945, Paris, France, 3 Institut National de la Sante´ et de la Recherche Me´dicale UMR S 945, Paris, France Abstract It has been proposed that ovale malaria in humans is caused by two closely related but distinct species of malaria parasites: P. ovale curtisi and P. ovale wallikeri. We have extended and optimized a Real-time PCR assay targeting the parasite’s small subunit ribosomal RNA (ssrRNA) gene to detect both these species. When the assay was applied to 31 archival blood samples from patients diagnosed with P. ovale, it was found that the infection in 20 was due to P. ovale curtisi and in the remaining 11 to P. ovale wallikeri. Thus, this assay provides a useful tool that can be applied to epidemiological investigations of the two newly recognized distinct P. ovale species, that might reveal if these species also differ in their clinical manifestation, drugs susceptibility and relapse periodicity. The results presented confirm that P. ovale wallikeri is not confined to Southeast Asia, since the majority of the patients analyzed in this study had acquired their P. ovale infection in African countries, mostly situated in West Africa. Citation: Calderaro A, Piccolo G, Gorrini C, Montecchini S, Rossi S, et al. (2012) A New Real-Time PCR for the Detection of Plasmodium ovale wallikeri. -

(PAD) and Post-Translational Protein Deimination—Novel Insights Into Alveolata Metabolism, Epigenetic Regulation and Host–Pathogen Interactions
biology Article Peptidylarginine Deiminase (PAD) and Post-Translational Protein Deimination—Novel Insights into Alveolata Metabolism, Epigenetic Regulation and Host–Pathogen Interactions Árni Kristmundsson 1,*, Ásthildur Erlingsdóttir 1 and Sigrun Lange 2,* 1 Institute for Experimental Pathology at Keldur, University of Iceland, Keldnavegur 3, 112 Reykjavik, Iceland; [email protected] 2 Tissue Architecture and Regeneration Research Group, School of Life Sciences, University of Westminster, London W1W 6UW, UK * Correspondence: [email protected] (Á.K.); [email protected] (S.L.) Simple Summary: Alveolates are a major group of free living and parasitic organisms; some of which are serious pathogens of animals and humans. Apicomplexans and chromerids are two phyla belonging to the alveolates. Apicomplexans are obligate intracellular parasites; that cannot complete their life cycle without exploiting a suitable host. Chromerids are mostly photoautotrophs as they can obtain energy from sunlight; and are considered ancestors of the apicomplexans. The pathogenicity and life cycle strategies differ significantly between parasitic alveolates; with some causing major losses in host populations while others seem harmless to the host. As the life cycles of Citation: Kristmundsson, Á.; Erlingsdóttir, Á.; Lange, S. some are still poorly understood, a better understanding of the factors which can affect the parasitic Peptidylarginine Deiminase (PAD) alveolates’ life cycles and survival is of great importance and may aid in new biomarker discovery. and Post-Translational Protein This study assessed new mechanisms relating to changes in protein structure and function (so-called Deimination—Novel Insights into “deimination” or “citrullination”) in two key parasites—an apicomplexan and a chromerid—to Alveolata Metabolism, Epigenetic assess the pathways affected by this protein modification. -

Highly Rearranged Mitochondrial Genome in Nycteria Parasites (Haemosporidia) from Bats
Highly rearranged mitochondrial genome in Nycteria parasites (Haemosporidia) from bats Gregory Karadjiana,1,2, Alexandre Hassaninb,1, Benjamin Saintpierrec, Guy-Crispin Gembu Tungalunad, Frederic Arieye, Francisco J. Ayalaf,3, Irene Landaua, and Linda Duvala,3 aUnité Molécules de Communication et Adaptation des Microorganismes (UMR 7245), Sorbonne Universités, Muséum National d’Histoire Naturelle, CNRS, CP52, 75005 Paris, France; bInstitut de Systématique, Evolution, Biodiversité (UMR 7205), Sorbonne Universités, Muséum National d’Histoire Naturelle, CNRS, Université Pierre et Marie Curie, CP51, 75005 Paris, France; cUnité de Génétique et Génomique des Insectes Vecteurs (CNRS URA3012), Département de Parasites et Insectes Vecteurs, Institut Pasteur, 75015 Paris, France; dFaculté des Sciences, Université de Kisangani, BP 2012 Kisangani, Democratic Republic of Congo; eLaboratoire de Biologie Cellulaire Comparative des Apicomplexes, Faculté de Médicine, Université Paris Descartes, Inserm U1016, CNRS UMR 8104, Cochin Institute, 75014 Paris, France; and fDepartment of Ecology and Evolutionary Biology, University of California, Irvine, CA 92697 Contributed by Francisco J. Ayala, July 6, 2016 (sent for review March 18, 2016; reviewed by Sargis Aghayan and Georges Snounou) Haemosporidia parasites have mostly and abundantly been de- and this lack of knowledge limits the understanding of the scribed using mitochondrial genes, and in particular cytochrome evolutionary history of Haemosporidia, in particular their b (cytb). Failure to amplify the mitochondrial cytb gene of Nycteria basal diversification. parasites isolated from Nycteridae bats has been recently reported. Nycteria parasites have been primarily described, based on Bats are hosts to a diverse and profuse array of Haemosporidia traditional taxonomy, in African insectivorous bats of two fami- parasites that remain largely unstudied. -

Desarrollo De Vacunas Frente a La Malaria
DESARROLLO DE VACUNAS FRENTE A LA MALARIA TRABAJO DE FIN DE GRADO Autor: Taranu, Alexandru Mirel Directores: Mesa Valle, Concepción María Garrido Cárdenas, José Antonio DEPARTAMENTO DE BIOLOGÍA Y GEOLOGÍA ÁREA DE PARASITOLOGÍA Grado en Biotecnología Junio, 2020 ÍNDICE 1. RESUMEN/ABSTRACT ..............................................................................................2 1.1. Resumen ......................................................................................................................2 1.2. Abstract .......................................................................................................................2 2. OBJETIVOS ...............................................................................................................3 3. INTRODUCCIÓN .......................................................................................................3 4. EL PARÁSITO ............................................................................................................5 4.1. Clasificación taxonómica .............................................................................................5 4.2. Ciclo de vida de Plasmodium .......................................................................................5 5. LA ENFERMEDAD .....................................................................................................6 5.1. Sintomatología ............................................................................................................6 5.2. Epidemiología ..............................................................................................................7 -

A Highly Rearranged Mitochondrial Genome in Nycteria Parasites
A highly rearranged mitochondrial genome in Nycteria parasites (Haemosporidia) from bats Gregory Karadjian, Alexandre Hassanin, Benjamin Saintpierre, Guy-Crispin Gembu Tungaluna, Frederic Ariey, Francisco J. Ayala, Irene Landau, Linda Duval To cite this version: Gregory Karadjian, Alexandre Hassanin, Benjamin Saintpierre, Guy-Crispin Gembu Tungaluna, Fred- eric Ariey, et al.. A highly rearranged mitochondrial genome in Nycteria parasites (Haemosporidia) from bats. Proceedings of the National Academy of Sciences of the United States of America , National Academy of Sciences, 2016, 113 (35), pp.9834 - 9839. 10.1073/pnas.1610643113. hal-01395176 HAL Id: hal-01395176 https://hal.sorbonne-universite.fr/hal-01395176 Submitted on 10 Nov 2016 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. Distributed under a Creative Commons Attribution - NonCommercial| 4.0 International License A highly rearranged mitochondrial genome in Nycteria parasites (Haemosporidia) from bats Gregory Karadjian a, b, 1, Alexandre Hassaninc, 1, Benjamin Saintpierred, Guy-Crispin Gembu -

Structural and Functional Insights Into Apicomplexan Gliding and Its Regulation
Structural and functional insights into apicomplexan gliding and its regulation Dissertation to obtain the degree of Doctor of Natural Sciences University of Hamburg Faculty of Mathematics, Informatics and Natural Sciences at the Department of Biology by Samuel Pažický from Bratislava, Slovakia Hamburg 2020 Examination commission Examination commission chair Prof. Dr. Jörg Ganzhorn (University of Hamburg) Examination commission members Prof. Jonas Schmidt-Chanasit (Bernhard Nocht Institute for Tropical Medicine and University of Hamburg) Prof. Tim Gilberger (Bernhard Nocht Institute for Tropical Medicine, Centre for Structural Systems Biology and University of Hamburg) Dr. Maria Garcia-Alai (European Molecular Biology Laboratory and Centre for Structural Systems Biology) Dr. Christian Löw (European Molecular Biology Laboratory and Centre for Structural Systems Biology) Date of defence: 29.01.2021 This work was performed at European Molecular Biology Laboratory, Hamburg Unit under the supervision of Dr. Christian Löw and Prof. Tim-Wolf Gilberger. The work was supported by the Joachim Herz Foundation. Evaluation Prof. Dr. rer. nat. Tim-Wolf Gilberger Bernhard Nocht Institute for Tropical Medicine (BNITM) Department of Cellular Parasitology Hamburg Dr. Christian Löw European Molecular Biology Laboratory Hamburg unit Hamburg Prof. Dr. vet. med. Thomas Krey Hannover Medical School Institute of Virology Declaration of academic honesty I hereby declare, on oath, that I have written the present dissertation by my own and have not used other than the acknowledged resources and aids. Eidesstattliche Erklärung Hiermit erkläre ich an Eides statt, dass ich die vorliegende Dissertationsschrift selbst verfasst und keine anderen als die angegebenen Quellen und Hilfsmittel benutzt habe. Hamburg, 22.9.2020 Samuel Pažický List of contents Declaration of academic honesty 4 List of contents 5 Acknowledgements 6 Summary 7 Zusammenfassung 10 List of publications 12 Scientific contribution to the manuscript 14 Abbreviations 16 1.