Indented Clado Exercise
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
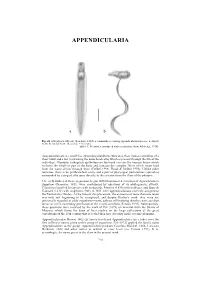
Appendicularia of CTAW
APPENDICULARIA APPENDICULARIA a b Fig. 26. Oikopleura albicans (Leuckart, 1854), a commonly occurring appendicularian species: a, dorsal view; b, lateral view. (Scale bar = 0.1 mm). [after T. Prentiss, reproduced with permission, from Alldredge 1976] Appendicularians are small free swimming planktonic tunicates, their bodies consisting of a short trunk and a tail (containing the notochord cells) which is present through the life of the individual. Glandular (oikoplast) epithelium on the trunk secretes the mucous house which encloses the whole or part of the body and contains the complex filters which strain food from the water driven through them (Deibel 1998; Flood & Deibel 1998). Unlike other tunicates, there is no peribranchial cavity and a pair of pharyngeal perforations (spiracles) surrounded by a ring of cilia open directly to the exterior from the floor of the pharynx. The early studies of these organisms, begun with Chamisso's description of Appendicularia flagellum Chamisso, 1821, were confounded by questions of its phylogenetic affinity. Chamisso classified his species with medusoids, Mertens (1830) with molluscs, and Quoy & Gaimard (1833) with zoophytes. Only in 1851 were appendicularians correctly assigned to the Tunicata by Huxley. At the time of this placement, the existence of more than one taxon was only just beginning to be recognised, and despite Huxley's work, they were not universally regarded as adult organisms—some authors still insisting that they were ascidian larvae or a free swimming generation of the sessile ascidians (Fenaux 1993). Subsequently, these questions were resolved by the work of Fol (1872) on material from the Straits of Messina, which forms the basis of later studies on the large collections of the great expeditions of the 19th century that revealed their true diversity in the oceanic plankton. -

From the Crato Formation (Lower Cretaceous)
ORYCTOS.Vol. 3 : 3 - 8. Décembre2000 FIRSTRECORD OT CALAMOPLEU RUS (ACTINOPTERYGII:HALECOMORPHI: AMIIDAE) FROMTHE CRATO FORMATION (LOWER CRETACEOUS) OF NORTH-EAST BRAZTL David M. MARTILL' and Paulo M. BRITO'z 'School of Earth, Environmentaland PhysicalSciences, University of Portsmouth,Portsmouth, POl 3QL UK. 2Departmentode Biologia Animal e Vegetal,Universidade do Estadode Rio de Janeiro, rua SâoFrancisco Xavier 524. Rio de Janeiro.Brazll. Abstract : A partial skeleton representsthe first occurrenceof the amiid (Actinopterygii: Halecomorphi: Amiidae) Calamopleurus from the Nova Olinda Member of the Crato Formation (Aptian) of north east Brazil. The new spe- cimen is further evidencethat the Crato Formation ichthyofauna is similar to that of the slightly younger Romualdo Member of the Santana Formation of the same sedimentary basin. The extended temporal range, ?Aptian to ?Cenomanian,for this genus rules out its usefulnessas a biostratigraphic indicator for the Araripe Basin. Key words: Amiidae, Calamopleurus,Early Cretaceous,Brazil Première mention de Calamopleurus (Actinopterygii: Halecomorphi: Amiidae) dans la Formation Crato (Crétacé inférieur), nord est du Brésil Résumé : la première mention dans le Membre Nova Olinda de la Formation Crato (Aptien ; nord-est du Brésil) de I'amiidé (Actinopterygii: Halecomorphi: Amiidae) Calamopleurus est basée sur la découverted'un squelettepar- tiel. Le nouveau spécimen est un élément supplémentaireindiquant que I'ichtyofaune de la Formation Crato est similaire à celle du Membre Romualdo de la Formation Santana, située dans le même bassin sédimentaire. L'extension temporelle de ce genre (?Aptien à ?Cénomanien)ne permet pas de le considérer comme un indicateur biostratigraphiquepour le bassin de l'Araripe. Mots clés : Amiidae, Calamopleurus, Crétacé inférieu4 Brésil INTRODUCTION Araripina and at Mina Pedra Branca, near Nova Olinda where cf. -

A Phylogenomic Analysis of Turtles ⇑ Nicholas G
Molecular Phylogenetics and Evolution 83 (2015) 250–257 Contents lists available at ScienceDirect Molecular Phylogenetics and Evolution journal homepage: www.elsevier.com/locate/ympev A phylogenomic analysis of turtles ⇑ Nicholas G. Crawford a,b,1, James F. Parham c, ,1, Anna B. Sellas a, Brant C. Faircloth d, Travis C. Glenn e, Theodore J. Papenfuss f, James B. Henderson a, Madison H. Hansen a,g, W. Brian Simison a a Center for Comparative Genomics, California Academy of Sciences, 55 Music Concourse Drive, San Francisco, CA 94118, USA b Department of Genetics, University of Pennsylvania, Philadelphia, PA 19104, USA c John D. Cooper Archaeological and Paleontological Center, Department of Geological Sciences, California State University, Fullerton, CA 92834, USA d Department of Biological Sciences, Louisiana State University, Baton Rouge, LA 70803, USA e Department of Environmental Health Science, University of Georgia, Athens, GA 30602, USA f Museum of Vertebrate Zoology, University of California, Berkeley, CA 94720, USA g Mathematical and Computational Biology Department, Harvey Mudd College, 301 Platt Boulevard, Claremont, CA 9171, USA article info abstract Article history: Molecular analyses of turtle relationships have overturned prevailing morphological hypotheses and Received 11 July 2014 prompted the development of a new taxonomy. Here we provide the first genome-scale analysis of turtle Revised 16 October 2014 phylogeny. We sequenced 2381 ultraconserved element (UCE) loci representing a total of 1,718,154 bp of Accepted 28 October 2014 aligned sequence. Our sampling includes 32 turtle taxa representing all 14 recognized turtle families and Available online 4 November 2014 an additional six outgroups. Maximum likelihood, Bayesian, and species tree methods produce a single resolved phylogeny. -

Eurochordata and Cephalochordata of Persian Gulf & Oman See
Subphylum Urochordata : Tunicates Class Ascidacea Class Larvacea Class Thaliacea Subphylum Cephalochordata : Amphioxus Phylum Chordata : 1 - Subphylum Urochordata 2 Subphylum Cephalochordata (Subphylum Urochordata : Tunicates ) Class Ascidiacea Class Larvacea Class Thaliacea (Subphylum Cephalochordata ) Class Ascidiacea Ascidia Kowalevsky Patricia Kott D. lane David George & Gordon Paterson Herdmania momus savigny Herdmania momus kiamanensis Didemnum candidum savigny Phallusia nigra savigny Styela canopus savigny Class Ascidiacea : Order : Aplousobranchia Family : Polyclinidae : Aplidium cf. rubripunctum C.Monniot & F. Monniot 1997 Polyclinum constellatum Savigny, 1816 Family Didemnidae Didemnum candidum Savigny, 1816 Didemnum cf. granulatum Tokioka, 1954 Didemnum obscurum F. Monniot 1969 Didemnum perlucidum F. Monniot, 1983 Didemnum sp. Didemnum yolky C.Monniot & F. Monniot 1997 Diplosoma listerianum Milne Edwards, 1841 Order : Phlebobranchia Family : Ascidiidae Ascidia sp. Phallusia julinea Sluiter, 1915 Phallusia nigra Savigny, 1816 Order : Stolidobranchia Family : Styelidae Botryllus gregalis Sluiter, 1898 Botryllus niger Herdman, 1886 Botryllus sp. Eusynstyela cf. hartmeyeri Michaelsen, 1904 Polyandrocarpa sp. Styela canopus Savigny, 1816 Symplegma bahraini C.Monniot & F. Monniot 1997 Symplegma brakenhielmi Michaelsen, 1904 Family pyuridae Herdmania momus Savigny, 1816 Pyura curvigona Tokioka, 1950 Pyurid sp. Class Larvacea Larva Seymour Sewell Phylum Chordata Subphylum Urochordata * Class Larvacea Order Copelata Family Fritillariidae -

The Origins of Chordate Larvae Donald I Williamson* Marine Biology, University of Liverpool, Liverpool L69 7ZB, United Kingdom
lopmen ve ta e l B Williamson, Cell Dev Biol 2012, 1:1 D io & l l o l g DOI: 10.4172/2168-9296.1000101 e y C Cell & Developmental Biology ISSN: 2168-9296 Research Article Open Access The Origins of Chordate Larvae Donald I Williamson* Marine Biology, University of Liverpool, Liverpool L69 7ZB, United Kingdom Abstract The larval transfer hypothesis states that larvae originated as adults in other taxa and their genomes were transferred by hybridization. It contests the view that larvae and corresponding adults evolved from common ancestors. The present paper reviews the life histories of chordates, and it interprets them in terms of the larval transfer hypothesis. It is the first paper to apply the hypothesis to craniates. I claim that the larvae of tunicates were acquired from adult larvaceans, the larvae of lampreys from adult cephalochordates, the larvae of lungfishes from adult craniate tadpoles, and the larvae of ray-finned fishes from other ray-finned fishes in different families. The occurrence of larvae in some fishes and their absence in others is correlated with reproductive behavior. Adult amphibians evolved from adult fishes, but larval amphibians did not evolve from either adult or larval fishes. I submit that [1] early amphibians had no larvae and that several families of urodeles and one subfamily of anurans have retained direct development, [2] the tadpole larvae of anurans and urodeles were acquired separately from different Mesozoic adult tadpoles, and [3] the post-tadpole larvae of salamanders were acquired from adults of other urodeles. Reptiles, birds and mammals probably evolved from amphibians that never acquired larvae. -

Towards an Understanding of Salp Swarm Dynamics. ICES CM 2002/N
CM 2002/ N:12 Towards an understanding of salp swarm dynamics by Patricia Kremer ABSTRACT Species of salps are characterized by intermittent blooms. Several studies have documented the importance of physical processes both in providing seed stocks of salps and creating an environment that is favorable for the rapid increase of salp populations. Although salps are typically oceanic, most observations of bloom dynamics have been made in more accessible inshore waters, so it is difficult to assess how frequent and widespread these swarms are. A review and comparison of existing data is helping to define geographic “hot spots” for salp blooms as well as the necessary physical and biological precursors. Although at this point, the review is far from complete, several generalities are emerging. The details of the physical forcing functions vary, but the overall physical regime seems to require a region of pulsed mixing of oceanic water that results in a relatively high standing stock of autotrophs. For a salp bloom to occur, there also needs to be an adequate seed population of salps and sufficient sustained primary production to support the biomass of the salp population as the bloom develops. As non-selective filter feeders, salps are able to remove a wide range of particulates from the water column, transforming the undigested portion into fast sinking feces. Therefore, when salps occur at high densities, the water is characterized by low abundance of other plankton, with obvious trophic implications. Patricia Kremer: Marine Sciences, University of Connecticut, Groton CT 06340-6097, USA [tel: +1 860 405 9140; fax: +1 860 405 9153; e-mail [email protected]] INTRODUCTION Salps are holoplanktonic grazers that have a life history, feeding biology, and population dynamics that contrasts sharply to copepods and other crustacean zooplankton (Madin and Deibel 1998). -

Evolution of the Iguanine Lizards (Sauria, Iguanidae) As Determined by Osteological and Myological Characters David F
Brigham Young University Science Bulletin, Biological Series Volume 12 | Number 3 Article 1 1-1971 Evolution of the iguanine lizards (Sauria, Iguanidae) as determined by osteological and myological characters David F. Avery Department of Biology, Southern Connecticut State College, New Haven, Connecticut Wilmer W. Tanner Department of Zoology, Brigham Young University, Provo, Utah Follow this and additional works at: https://scholarsarchive.byu.edu/byuscib Part of the Anatomy Commons, Botany Commons, Physiology Commons, and the Zoology Commons Recommended Citation Avery, David F. and Tanner, Wilmer W. (1971) "Evolution of the iguanine lizards (Sauria, Iguanidae) as determined by osteological and myological characters," Brigham Young University Science Bulletin, Biological Series: Vol. 12 : No. 3 , Article 1. Available at: https://scholarsarchive.byu.edu/byuscib/vol12/iss3/1 This Article is brought to you for free and open access by the Western North American Naturalist Publications at BYU ScholarsArchive. It has been accepted for inclusion in Brigham Young University Science Bulletin, Biological Series by an authorized editor of BYU ScholarsArchive. For more information, please contact [email protected], [email protected]. S-^' Brigham Young University f?!AR12j97d Science Bulletin \ EVOLUTION OF THE IGUANINE LIZARDS (SAURIA, IGUANIDAE) AS DETERMINED BY OSTEOLOGICAL AND MYOLOGICAL CHARACTERS by David F. Avery and Wilmer W. Tanner BIOLOGICAL SERIES — VOLUME Xil, NUMBER 3 JANUARY 1971 Brigham Young University Science Bulletin -

A Small Lepidosauromorph Reptile from the Early Triassic of Poland
A SMALL LEPIDOSAUROMORPH REPTILE FROM THE EARLY TRIASSIC OF POLAND SUSAN E. EVANS and MAGDALENA BORSUK−BIAŁYNICKA Evans, S.E. and Borsuk−Białynicka, M. 2009. A small lepidosauromorph reptile from the Early Triassic of Poland. Palaeontologia Polonica 65, 179–202. The Early Triassic karst deposits of Czatkowice quarry near Kraków, southern Poland, has yielded a diversity of fish, amphibians and small reptiles. Two of these reptiles are lepido− sauromorphs, a group otherwise very poorly represented in the Triassic record. The smaller of them, Sophineta cracoviensis gen. et sp. n., is described here. In Sophineta the unspecial− ised vertebral column is associated with the fairly derived skull structure, including the tall facial process of the maxilla, reduced lacrimal, and pleurodonty, that all resemble those of early crown−group lepidosaurs rather then stem−taxa. Cladistic analysis places this new ge− nus as the sister group of Lepidosauria, displacing the relictual Middle Jurassic genus Marmoretta and bringing the origins of Lepidosauria closer to a realistic time frame. Key words: Reptilia, Lepidosauria, Triassic, phylogeny, Czatkowice, Poland. Susan E. Evans [[email protected]], Department of Cell and Developmental Biology, Uni− versity College London, Gower Street, London, WC1E 6BT, UK. Magdalena Borsuk−Białynicka [[email protected]], Institut Paleobiologii PAN, Twarda 51/55, PL−00−818 Warszawa, Poland. Received 8 March 2006, accepted 9 January 2007 180 SUSAN E. EVANS and MAGDALENA BORSUK−BIAŁYNICKA INTRODUCTION Amongst living reptiles, lepidosaurs (snakes, lizards, amphisbaenians, and tuatara) form the largest and most successful group with more than 7 000 widely distributed species. The two main lepidosaurian clades are Rhynchocephalia (the living Sphenodon and its extinct relatives) and Squamata (lizards, snakes and amphisbaenians). -

Tiago Rodrigues Simões
Diapsid Phylogeny and the Origin and Early Evolution of Squamates by Tiago Rodrigues Simões A thesis submitted in partial fulfillment of the requirements for the degree of Doctor of Philosophy in SYSTEMATICS AND EVOLUTION Department of Biological Sciences University of Alberta © Tiago Rodrigues Simões, 2018 ABSTRACT Squamate reptiles comprise over 10,000 living species and hundreds of fossil species of lizards, snakes and amphisbaenians, with their origins dating back at least as far back as the Middle Jurassic. Despite this enormous diversity and a long evolutionary history, numerous fundamental questions remain to be answered regarding the early evolution and origin of this major clade of tetrapods. Such long-standing issues include identifying the oldest fossil squamate, when exactly did squamates originate, and why morphological and molecular analyses of squamate evolution have strong disagreements on fundamental aspects of the squamate tree of life. Additionally, despite much debate, there is no existing consensus over the composition of the Lepidosauromorpha (the clade that includes squamates and their sister taxon, the Rhynchocephalia), making the squamate origin problem part of a broader and more complex reptile phylogeny issue. In this thesis, I provide a series of taxonomic, phylogenetic, biogeographic and morpho-functional contributions to shed light on these problems. I describe a new taxon that overwhelms previous hypothesis of iguanian biogeography and evolution in Gondwana (Gueragama sulamericana). I re-describe and assess the functional morphology of some of the oldest known articulated lizards in the world (Eichstaettisaurus schroederi and Ardeosaurus digitatellus), providing clues to the ancestry of geckoes, and the early evolution of their scansorial behaviour. -

Clades™ Prehistoric Card Game a Clade Is a Section of the Evolutionary Family Tree—Basically Any Branch, Including All Its Sub-Branches
CLADES™ PREHISTORIC Card Game A clade is a section of the evolutionary family tree —basically any branch, including all its sub-branches. A clade is a family of organisms, or living things, that are all more closely related to each other than they are to any other organisms. In this game you match cards according to their clades. Contents: Deck of 83 Clades Prehistoric cards. Includes 27 cards of each color and 2 bonus cards. There are also 5 animal description cards not used in play. Object: Spot matching card triples to collect the biggest animal pile! Setup Deal 1 face-down card to each player as their personal card. For now, players keep these cards face-down and don’t look at them. Deal 12 face-down shared cards to the middle of the play area. If you’re learning or teaching the game: • Before dealing, set aside the bonus cards and the cards showing only one or two animals. Play with just the cards showing three animals. • Deal 7 shared cards instead of 12. CLADESPrehistoricRules2.indd 1 10/17/17 10:15 AM All players help flip the 12 shared cards face-up. Sort the cards into three Making Triples rows according to their clades: top for Mammalia (mammals), middle for Sauropsida (sauropsids, or reptiles and birds), and bottom for Arthropoda In Clades Prehistoric, any two cards can make a triple with exactly one other (arthropods, or “bugs”). When the table is ready, each player picks up their card in the deck. personal card and looks at it. -

Antimicrobial Peptides in Reptiles
Pharmaceuticals 2014, 7, 723-753; doi:10.3390/ph7060723 OPEN ACCESS pharmaceuticals ISSN 1424-8247 www.mdpi.com/journal/pharmaceuticals Review Antimicrobial Peptides in Reptiles Monique L. van Hoek National Center for Biodefense and Infectious Diseases, and School of Systems Biology, George Mason University, MS1H8, 10910 University Blvd, Manassas, VA 20110, USA; E-Mail: [email protected]; Tel.: +1-703-993-4273; Fax: +1-703-993-7019. Received: 6 March 2014; in revised form: 9 May 2014 / Accepted: 12 May 2014 / Published: 10 June 2014 Abstract: Reptiles are among the oldest known amniotes and are highly diverse in their morphology and ecological niches. These animals have an evolutionarily ancient innate-immune system that is of great interest to scientists trying to identify new and useful antimicrobial peptides. Significant work in the last decade in the fields of biochemistry, proteomics and genomics has begun to reveal the complexity of reptilian antimicrobial peptides. Here, the current knowledge about antimicrobial peptides in reptiles is reviewed, with specific examples in each of the four orders: Testudines (turtles and tortosises), Sphenodontia (tuataras), Squamata (snakes and lizards), and Crocodilia (crocodilans). Examples are presented of the major classes of antimicrobial peptides expressed by reptiles including defensins, cathelicidins, liver-expressed peptides (hepcidin and LEAP-2), lysozyme, crotamine, and others. Some of these peptides have been identified and tested for their antibacterial or antiviral activity; others are only predicted as possible genes from genomic sequencing. Bioinformatic analysis of the reptile genomes is presented, revealing many predicted candidate antimicrobial peptides genes across this diverse class. The study of how these ancient creatures use antimicrobial peptides within their innate immune systems may reveal new understandings of our mammalian innate immune system and may also provide new and powerful antimicrobial peptides as scaffolds for potential therapeutic development. -

History of Fishes - Structural Patterns and Trends in Diversification
History of fishes - Structural Patterns and Trends in Diversification AGNATHANS = Jawless • Class – Pteraspidomorphi • Class – Myxini?? (living) • Class – Cephalaspidomorphi – Osteostraci – Anaspidiformes – Petromyzontiformes (living) Major Groups of Agnathans • 1. Osteostracida 2. Anaspida 3. Pteraspidomorphida • Hagfish and Lamprey = traditionally together in cyclostomata Jaws = GNATHOSTOMES • Gnathostomes: the jawed fishes -good evidence for gnathostome monophyly. • 4 major groups of jawed vertebrates: Extinct Acanthodii and Placodermi (know) Living Chondrichthyes and Osteichthyes • Living Chondrichthyans - usually divided into Selachii or Elasmobranchi (sharks and rays) and Holocephali (chimeroids). • • Living Osteichthyans commonly regarded as forming two major groups ‑ – Actinopterygii – Ray finned fish – Sarcopterygii (coelacanths, lungfish, Tetrapods). • SARCOPTERYGII = Coelacanths + (Dipnoi = Lung-fish) + Rhipidistian (Osteolepimorphi) = Tetrapod Ancestors (Eusthenopteron) Close to tetrapods Lungfish - Dipnoi • Three genera, Africa+Australian+South American ACTINOPTERYGII Bichirs – Cladistia = POLYPTERIFORMES Notable exception = Cladistia – Polypterus (bichirs) - Represented by 10 FW species - tropical Africa and one species - Erpetoichthys calabaricus – reedfish. Highly aberrant Cladistia - numerous uniquely derived features – long, independent evolution: – Strange dorsal finlets, Series spiracular ossicles, Peculiar urohyal bone and parasphenoid • But retain # primitive Actinopterygian features = heavy ganoid scales (external