Impact of Invasive Tree Species on Natural Regeneration Species Composition, Diversity, and Density
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Dr. Duke's Phytochemical and Ethnobotanical Databases List of Plants for Tinnitus
Dr. Duke's Phytochemical and Ethnobotanical Databases List of Plants for Tinnitus Plant Chemical Count Activity Count Newcastelia viscida 1 1 Platanus occidentalis 1 1 Tacca aspera 1 1 Avicennia tomentosa 2 1 Coccoloba excoriata 1 1 Diospyros morrisiana 1 1 Cassia siamea 1 1 Diospyros derra 1 1 Rhododendron ledebourii 1 1 Thymelaea hirsuta 1 1 Dichrostachys glomerata 1 1 Diospyros wallichii 2 1 Erythroxylum gracilipes 1 1 Hyptis emoryi 1 1 Lemaireocereus thurberi 1 1 Pongamia pinnata 1 1 Quercus championi 2 1 Rubus spectabilis 2 1 Tetracera scandens 2 1 Arbutus menziesii 1 1 Betula sp. 2 1 Dillenia pentagyna 2 1 Erythroxylum rotundifolium 1 1 Grewia tiliaefolia 1 1 Inga punctata 1 1 Lepechinia hastata 1 1 Paeonia japonica 1 1 Plant Chemical Count Activity Count Pouteria torta 1 1 Rabdosia adenantha 1 1 Selaginella delicatula 1 1 Stemonoporus affinis 2 1 Rosa davurica 1 1 Calophyllum lankaensis 1 1 Colubrina granulosa 1 1 Acrotrema uniflorum 1 1 Diospyros hirsuta 2 1 Pedicularis palustris 1 1 Pistacia major 1 1 Psychotria adenophylla 2 1 Buxus microphylla 2 1 Clinopodium umbrosum 1 1 Diospyros maingayi 2 1 Epilobium rosmarinifolium 1 1 Garcinia xanthochymus 1 1 Hippuris vulgare 1 1 Kleinhovia hospita 1 1 Crotalaria semperflorens 1 1 Diospyros abyssinica 2 1 Isodon grandifolius 1 1 Salvia mexicana 1 1 Shorea affinis 2 1 Diospyros singaporensis 2 1 Erythroxylum amazonicum 1 1 Euclea crispa 1 1 2 Plant Chemical Count Activity Count Givotia rottleriformis 2 1 Zizyphus trinervia 2 1 Simaba obovata 1 1 Betula cordifolia 1 1 Platanus orientalis 1 1 Triadenum japonicum 1 1 Woodfordia floribunda 2 1 Calea zacatechichi 1 1 Diospyros natalensis 1 1 Alyxia buxifolia 1 1 Brassica napus var. -

REU Mentors and Projects 2018 (Updated January 11, 2018) UMBS
REU Mentors and Projects 2018 (updated January 11, 2018) UMBS forest ecosystem study Chris Gough, Virginia Commonwealth University Luke Nave, University of Michigan Chris Vogel, University of Michigan Forests of northern Michigan provide ecosystem services including the capture and sequestration of carbon, retention of nutrients, maintenance of organismal and ecosystem diversity, and protection of surface and ground water quality. Our collaborative team conducts research on the scientific underpinnings of these ecosystem services, with particular emphasis on plant and ecosystem ecology, disturbance ecology, ecological succession, carbon and nitrogen biogeochemistry, botany and mycology. We conduct this research in a variety of settings, including a 20-year experimental forest with a long-running carbon "flux" tower, a landscape- scale experimental disturbance in which >6700 trees were stem girdled, a pair of long-term chronosequences with stands from 20 to >200 years old, and, beginning in 2018, a new experimental manipulation of disturbance severity in which tree mortality will range from 45 to 85 %. REU student collaborators on the UMBS Forest Ecosystem Study team have numerous research options. Some examples include: 1) disturbance, climate, and forest age effects on carbon cycling; 2) mechanisms sustaining high rates of carbon storage in old forests; 3) remotely sensed ecosystem structure-carbon cycling relationships; and 4) fungal processes controlling decomposition and tree nutrient supply. Effects of climate change on wetland biodiversity Amy Schrank, University of Minnesota Climate change is predicted to result in significant losses in both the amount of and the biodiversity within Great Lakes coastal wetlands. Coastal wetlands provide important ecosystem services including nursery areas for the majority of Great Lakes fish species, important habitat for wildlife including rare and endangered species, a filter for pollutants and sediment, shoreline protection against wind and waves, and many others. -
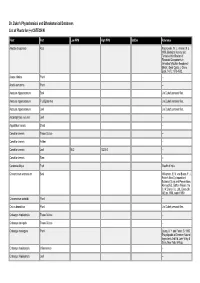
Dr. Duke's Phytochemical and Ethnobotanical Databases List of Plants for (+)-CATECHIN
Dr. Duke's Phytochemical and Ethnobotanical Databases List of Plants for (+)-CATECHIN Plant Part Low PPM High PPM StdDev Reference Abutilon theophrasti Root Paszkowski, W. L., Kremer, R. J. 1988. Biological Activity and Tentative Identification of Flavonoid Components in Velvetleaf (Abutilon theophrasti Medik.) Seed Coats. J. Chem. Ecol., 14(7): 1573-1582. Acacia nilotica Plant -- Acacia decurrens Plant -- Aesculus hippocastanum Bark Jim Duke's personal files. Aesculus hippocastanum Fruit Epidermis Jim Duke's personal files. Aesculus hippocastanum Leaf Jim Duke's personal files. Arctostaphylos uva-ursi Leaf -- Aspalathus linearis Shoot -- Camellia sinensis Tissue Culture -- Camellia sinensis Anther -- Camellia sinensis Leaf 85.2 13200.0 -- Camellia sinensis Stem -- Ceratonia siliqua Fruit Wealth of India. Cinnamomum aromaticum Bark Williamson, E. M. and Evans, F. J., Potter's New Cyclopaedia of Botanical Drugs and Preparations, Revised Ed., Saffron Walden, the C. W. Daniel Co., Ltd., Essex UK, 362 pp, 1988, reprint 1989. Cinnamomum sieboldii Plant -- Cnicus benedictus Plant Jim Duke's personal files. Crataegus rhipidophylla Tissue Culture -- Crataegus laevigata Tissue Culture -- Crataegus monogyna Plant Leung, A. Y. and Foster, S. 1995. Encyclopedia of Common Natural Ingredients 2nd Ed. John Wiley & Sons, New York. 649 pp. Crataegus rhipidophylla Inflorescence -- Crataegus rhipidophylla Leaf -- Plant Part Low PPM High PPM StdDev Reference Crataegus laevigata Leaf -- Crataegus laevigata Plant Leung, A. Y. and Foster, S. 1995. Encyclopedia of Common Natural Ingredients 2nd Ed. John Wiley & Sons, New York. 649 pp. Crataegus rhipidophylla Bud -- Crataegus rhipidophylla Flower -- Crataegus laevigata Bud -- Crataegus laevigata Flower -- Croton lechleri Plant -- Eucalyptus globulus Leaf -- Fagopyrum esculentum Root -- Fagopyrum esculentum Tissue Culture -- Fallopia japonica Root 2.0 -- Geranium thunbergii Tissue Culture -- Ginkgo biloba Tissue Culture Jim Duke's personal files. -

Tree Species Associations of Pinus Echinata Mill. Over a Large-Scale Sampling Regime on the Interior Highlands of Arkansas
TREE SPECIES ASSOCIATIONS OF PINUS ECHINATA MILL. OVER A LARGE-SCALE SAMPLING REGIME ON THE INTERIOR HIGHLANDS OF ARKANSAS James F. Rosson, Jr.1 ABSTRACT.—The Interior Highlands physiographic province of Arkansas is considered the ecological center of the geographic distribution of shortleaf pine (Pinus echinata Mill.). I used data from the U.S. Forest Service, Forest Inventory and Analysis (FIA) program to identify the major tree species associates of P. echinata across this 66,700-km2 landscape. Across the region, 41,207 km2 were covered by timberland. The study population was represented by 434 relatively undisturbed upland sample plots from the 1995 forest survey of Arkansas. P. echinata ≥12.7 cm in diameter at breast height (DBH) occurred on 211 of these sample plots. Additionally, it ranked fi rst in basal area on 119 plots, second on 39 plots, and third on 19 plots. Where P. echinata was dominant, stand basal area averaged 23.1 m2 ha-1 (± 0.57 SEM). I used chi-square to test for degree of association between the stand dominants and to test for positive and negative associations. There was a positive association between P. echinata and Quercus alba L. (χ2 = 0.490; 1df). In contrast, there was a negative association between P. echinata and Q. velutina Lam. (χ2 = 15.571; 1df). These results demonstrate that the chi- square test of association is effective even on the larger scales of sampling where lack of sample homogeneity may sometimes complicate analysis. Such quantitative tests for species associations offer meaningful insights into P. echinata communities at the landscape scale of sampling. -

Are the Consequences If Some Algal Species Are Lost? Saarinen, Anniina; Salovius-Lauren, Sonja; Mattila, Johanna
This is an electronic reprint of the original article. This reprint may differ from the original in pagination and typographic detail. Epifaunal community composition in five macroalgal species - What are the consequences if some algal species are lost? Saarinen, Anniina; Salovius-Lauren, Sonja; Mattila, Johanna Published in: Estuarine, Coastal and Shelf Science DOI: 10.1016/j.ecss.2017.08.009 Publicerad: 01/01/2018 Document Version (Referentgranskad version om publikationen är vetenskaplig) Document License CC BY-NC-ND Link to publication Please cite the original version: Saarinen, A., Salovius-Lauren, S., & Mattila, J. (2018). Epifaunal community composition in five macroalgal species - What are the consequences if some algal species are lost? Estuarine, Coastal and Shelf Science, 207, 402–413. https://doi.org/10.1016/j.ecss.2017.08.009 General rights Copyright and moral rights for the publications made accessible in the public portal are retained by the authors and/or other copyright owners and it is a condition of accessing publications that users recognise and abide by the legal requirements associated with these rights. Take down policy If you believe that this document breaches copyright please contact us providing details, and we will remove access to the work immediately and investigate your claim. This document is downloaded from the Research Information Portal of ÅAU: 11. Oct. 2021 1 1 Epifaunal community composition in five macroalgal species – what 2 are the consequences if some algal species are lost? 3 1 4 Corresponding -

Dr. Duke's Phytochemical and Ethnobotanical Databases List of Plants for Lyme Disease (Chronic)
Dr. Duke's Phytochemical and Ethnobotanical Databases List of Plants for Lyme Disease (Chronic) Plant Chemical Count Activity Count Garcinia xanthochymus 1 1 Nicotiana rustica 1 1 Acacia modesta 1 1 Galanthus nivalis 1 1 Dryopteris marginalis 2 1 Premna integrifolia 1 1 Senecio alpinus 1 1 Cephalotaxus harringtonii 1 1 Comptonia peregrina 1 1 Diospyros rotundifolia 1 1 Alnus crispa 1 1 Haplophyton cimicidum 1 1 Diospyros undulata 1 1 Roylea elegans 1 1 Bruguiera gymnorrhiza 1 1 Gmelina arborea 1 1 Orthosphenia mexicana 1 1 Lumnitzera racemosa 1 1 Melilotus alba 2 1 Duboisia leichhardtii 1 1 Erythroxylum zambesiacum 1 1 Salvia beckeri 1 1 Cephalotaxus spp 1 1 Taxus cuspidata 3 1 Suaeda maritima 1 1 Rhizophora mucronata 1 1 Streblus asper 1 1 Plant Chemical Count Activity Count Dianthus sp. 1 1 Glechoma hirsuta 1 1 Phyllanthus flexuosus 1 1 Euphorbia broteri 1 1 Hyssopus ferganensis 1 1 Lemaireocereus thurberi 1 1 Holacantha emoryi 1 1 Casearia arborea 1 1 Fagonia cretica 1 1 Cephalotaxus wilsoniana 1 1 Hydnocarpus anthelminticus 2 1 Taxus sp 2 1 Zataria multiflora 1 1 Acinos thymoides 1 1 Ambrosia artemisiifolia 1 1 Rhododendron schotense 1 1 Sweetia panamensis 1 1 Thymelaea hirsuta 1 1 Argyreia nervosa 1 1 Carapa guianensis 1 1 Parthenium hysterophorus 1 1 Rhododendron anthopogon 1 1 Strobilanthes cusia 1 1 Dianthus superbus 1 1 Pyropolyporus fomentarius 1 1 Euphorbia hermentiana 1 1 Porteresia coarctata 1 1 2 Plant Chemical Count Activity Count Aerva lanata 1 1 Rivea corymbosa 1 1 Solanum mammosum 1 1 Juniperus horizontalis 1 1 Maytenus -

Mercury Bioaccumulation in Some Commercially Valuable Marine Organisms from Mosa Bay, Persian Gulf
Int. J. Environ. Res., 5(3):757-762, Summer 2011 ISSN: 1735-6865 Mercury Bioaccumulation in Some Commercially Valuable Marine Organisms from Mosa Bay, Persian Gulf * Mortazavi, M. S. and Sharifian, S. Persian Gulf and Oman Sea Ecological Research Institute, P. O. Box 79145-1597 Bandar Abbas, Hormozgan, Iran Received 23 Sep. 2010; Revised 11 March 2011; Accepted 17 March 2011 ABSTRACT: This study was undertaken to determine the concentration of mercury in edible muscle of five commercially valuable marine organisms from Mosa Bay, Persian Gulf, Iran. The total mercury concentrations were determined by cold vapour atomic absorption spectrophotometry and expressed in µg/g dry weight. Certified reference materials were used for each batch of analysis. Mercury concentration was 0.373 µg/g for Liza abu, 1.172 µg/g Sparidentex hasta, 0.445µg/g for Acanthopagrus latus, 0.390 µg/g for Thunnus tonggol, and 0.360 µg/g for Fenneropenaeus indicus. Carnivorous fish had higher level of mercury than non-carnivorous. Statistical analysis revealed weak correlation between fish mercury concentration and length for all studied organisms. The observed concentrations were comparable to those found in other areas of the Persian Gulf and were lower than the WHO guideline of 0.5 mg/kg wet weight. Our result demonstrated that estimated daily and weekly intakes of mercury via consumption of fish flesh were far below the PTDI and PTWI values recommended by FAO/WHO. Key words: Mercury,Marine Organisms, Food Safety, Persian Gulf, Mosa Bay INTRODUCTION Fish is widely consumed in many parts of the world Bay disaster in Japan, which has resulted in the deaths by humans because it has high protein content, low of over 1,000 people and left over 2,000 seriously ill saturated fat and also contains omega fatty acids from contaminated seafood (Harada 1995). -

Do Arthropod Assemblages Fit the Grassland and Savanna Biomes Of
Research Article Arthropod assemblages of grassland and savanna in South Africa Page 1 of 10 Do arthropod assemblages fit the grassland and AUTHORS: savanna biomes of South Africa? Monique Botha1 Stefan J. Siebert1 Johnnie van den Berg1 The long-standing tradition of classifying South Africa’s biogeographical area into biomes is commonly linked to vegetation structure and climate. Because arthropod communities are often governed by both AFFILIATION: these factors, it can be expected that arthropod communities would fit the biomes. To test this hypothesis, 1Unit for Environmental Sciences we considered how well arthropod species assemblages fit South Africa’s grassy biomes. Arthropod and Management, North-West assemblages were sampled from six localities across the grassland and savanna biomes by means University, Potchefstroom, South Africa of suction sampling, to determine whether the two biomes have distinctive arthropod assemblages. Arthropod samples of these biomes clustered separately in multidimensional scaling analyses. Within CORRESPONDENCE TO: biomes, arthropod assemblages were more distinctive for savanna localities than grassland. Arthropod Monique Botha samples of the two biomes clustered together when trophic groups were considered separately, suggesting some similarity in functional assemblages. Dissimilarity was greatest between biomes for EMAIL: phytophagous and predacious trophic groups, with most pronounced differentiation between biomes [email protected] at sub-escarpment localities. Our results indicate that different arthropod assemblages do fit the grassy DATES: biomes to some extent, but the pattern is not as clear as it is for plant species. Received: 12 Nov. 2015 Significance: Revised: 26 Mar. 2016 • Provides the first comparison of arthropod composition between grassland and savanna biomes of Accepted: 06 Apr. -

Advantageous Extraction, Cleanup, and UHPLC-MS/MS Detection of Patulin Mycotoxin in Dietary Supplements and Herbal Blends Containing Hawberry from Crataegus Spp
Hindawi Journal of Analytical Methods in Chemistry Volume 2019, Article ID 2159097, 13 pages https://doi.org/10.1155/2019/2159097 Research Article Advantageous Extraction, Cleanup, and UHPLC-MS/MS Detection of Patulin Mycotoxin in Dietary Supplements and Herbal Blends Containing Hawberry from Crataegus spp. Anna Przybylska ,1 Grzegorz Bazylak ,1 Robert Kosicki,2 Iwona Altyn ,2 Magdalena Twaruzek ,2 Jan Grajewski ,2 and Anna Soltys-Lelek3 1Department of Pharmaco-Bromatology and Molecular Nutrition, Faculty of Pharmacy, Collegium Medicum, Nicolaus Copernicus University, Jagiellonska 13, PL-85067 Bydgoszcz, Poland 2Department of Physiology and Toxicology, Institute of Experimental Biology, Faculty of Natural Sciences, Kazimierz Wielki University, Chodkiewicza 30, PL-85064 Bydgoszcz, Poland 3Ojcow National Park, Ojcow 9, PL-32045 Suloszowa, Poland Correspondence should be addressed to Grzegorz Bazylak; [email protected] Received 14 October 2018; Revised 21 December 2018; Accepted 13 January 2019; Published 6 February 2019 Academic Editor: Nu´ria Fontanals Copyright © 2019 Anna Przybylska et al. /is is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. Patulin (PAT) is a highly genotoxic mycotoxin still found as the common contaminant of various kinds of spoiled fruits and related commodities which are often endorsed as the health-enhancing products. /us, a fast and convenient liquid-solid extraction followed by a solid-phase cleanup with the MycoSep 228 AflaPat multifunctional column was used for the highly efficient isolation of PAT with an average recovery of 112.7% from® commercial dietary supplements and herbal blends formulated with dried hawberry. -

An Introduction to Evolutionary Ecology
An Introduction to Evolutionary Ecology ANDREW COCKBURN Professor of Botany and Zoology School of Life Sciences Australian National University Canberra, Australia ILLUSTRATED BY KARINA HANSEN OXFORD BLACKWELL SCIENTIFIC PUBLICATIONS LONDON EDINBURGH BOSTON MELBOURNE PARIS BERLIN VIENNA Contents PREFACE, xi THE SCOPE OF EVOLUTIONARY ECOLOGY, 1 Patterns in the living world, 2 Why was 1983 so bad for giant pandas?, 2 Different ways of growing up, 3 Why have sex in the sea?, 6 Why does each fig have its own fig wasp?, 7 Interactions between species: a periodic table?, 11 Why are there so many species of beetles?, 12 Darwin's solution to asking why?, 13 Evolution is not a synonym of natural selection, 14 What is natural selection?, 14 Types of selection, 15 Directional, stabilising and disruptive selection, 16 Frequency-dependent selection, 16 Density-dependent and density-independent selection, 18 Hard and soft selection, 19 Natural and artificial selection, 19 Natural and sexual selection, 19 What are fitness and adaptation?, 20 The theory of natural selection is not tautologous, 20 Absolute versus relative prowess, 21 How'do we detect selection and adaptation?, 22 Summary, 23 Further reading, 23 Topics for discussion, 24 THE GENETIC BASIS OF EVOLUTIONARY CHANGE, 25 Genotype and phenotype, 26 Alleles of genes, and dominance, 28 Gene interactions, 28 Nature versus nurture and heritability, 29 How does genetic variation arise?, 32 How do allele frequencies change?, 33 The Hardy—Weinberg Law, 33 Random causes, 34 The Neutral Theory of Molecular -

REU Mentors and Projects 2018 (Updated January 11, 2018) UMBS
REU Mentors and Projects 2018 (updated January 11, 2018) UMBS forest ecosystem study Chris Gough, Virginia Commonwealth University Luke Nave, University of Michigan Chris Vogel, University of Michigan Forests of northern Michigan provide ecosystem services including the capture and sequestration of carbon, retention of nutrients, maintenance of organismal and ecosystem diversity, and protection of surface and ground water quality. Our collaborative team conducts research on the scientific underpinnings of these ecosystem services, with particular emphasis on plant and ecosystem ecology, disturbance ecology, ecological succession, carbon and nitrogen biogeochemistry, botany and mycology. We conduct this research in a variety of settings, including a 20-year experimental forest with a long-running carbon "flux" tower, a landscape- scale experimental disturbance in which >6700 trees were stem girdled, a pair of long-term chronosequences with stands from 20 to >200 years old, and, beginning in 2018, a new experimental manipulation of disturbance severity in which tree mortality will range from 45 to 85 %. REU student collaborators on the UMBS Forest Ecosystem Study team have numerous research options. Some examples include: 1) disturbance, climate, and forest age effects on carbon cycling; 2) mechanisms sustaining high rates of carbon storage in old forests; 3) remotely sensed ecosystem structure-carbon cycling relationships; and 4) fungal processes controlling decomposition and tree nutrient supply. Effects of climate change on wetland biodiversity Amy Schrank, University of Minnesota Climate change is predicted to result in significant losses in both the amount of and the biodiversity within Great Lakes coastal wetlands. Coastal wetlands provide important ecosystem services including nursery areas for the majority of Great Lakes fish species, important habitat for wildlife including rare and endangered species, a filter for pollutants and sediment, shoreline protection against wind and waves, and many others. -

Javaslatok a NATURA2000 Jelölőfajok Bővítésére – Közösségi Jelentőségű Rózsa- És Galagonyafajok
GYEPGAZDÁLKODÁSI KÖZLEMÉNYEK , 2015 (1-2). Javaslatok a NATURA2000 jelöl őfajok bővítésére – közösségi jelent őség ű rózsa- és galagonyafajok Kerényi-Nagy Viktor – Penksza Károly Szent István Egyetem Növénytani és Ökofiziológiai Intézet, Növénytani Tanszék, Gödöll ő [email protected] ÖSSZEFOGLALÁS species can be found in the territory of the European Union. Furthermore these species grow in natural areas or localities Szerbia csatlakozása révén lehet őség nyílik a Natura 2000 which are hardly disturbed by human activities, therefore jelöl őfajok listájának kiegészítésére, b ővítésére. A rózsafélék protecting them is important for nature conservation and (Rosaceae) családjának rendkívül összetett taxonómiai és community interrest as well. nevezéktani nehézségei lehetnek az okai annak, hogy a család fajai We have presented in detail the protecting category, the csak igen visszafogottan képviselik magukat a Natura 2000 morphology, the blooming time, the mistakable species, the habitat jelöl őfajok listáján. Az alábbi fajok populációinak dönt ő többsége and the size of the population, the possible dangers, the habitat- az Európai Unió területére, azon belül is a Pannon Biogeográfiai treatment and the species-protection of 5 roses (R. zalana, R. Régióba esik, ezen fajok kimerítik a Natura 2000 kritériumát, pocsii, R. hungarica, R. gizellae and R. stylosa) and 4 hawthorns természetes vagy természet közeli term őhelyek jelz ői, így védelm űk (C. nigra, C. ovalis, C. rosaeformis, C. lindmanii). a természetvédelmi kérdéseken felül még közösségi érdek is. For the easier recognition of the species, the sect. Részletesen bemutattuk az 5 rózsa (Rosa zalana, Rosa pocsii, Rubiginosae are compared on the 1st chart. pictures of all the Rosa hungarica, Rosa gizellae és Rosa stylosa) és 4 galagonya species are shown, their habitat in Europe and in Hungary or in (Crataegus nigra, Crataegus ovalis, Crataegus rosaeformis, the Carpathian-basin is presented as well.