10.2 Performance of Planted Native Conifer Trees MEASUREMENT of STANDS
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Supporting Information
Supporting Information Mao et al. 10.1073/pnas.1114319109 SI Text BEAST Analyses. In addition to a BEAST analysis that used uniform Selection of Fossil Taxa and Their Phylogenetic Positions. The in- prior distributions for all calibrations (run 1; 144-taxon dataset, tegration of fossil calibrations is the most critical step in molecular calibrations as in Table S4), we performed eight additional dating (1, 2). We only used the fossil taxa with ovulate cones that analyses to explore factors affecting estimates of divergence could be assigned unambiguously to the extant groups (Table S4). time (Fig. S3). The exact phylogenetic position of fossils used to calibrate the First, to test the effect of calibration point P, which is close to molecular clocks was determined using the total-evidence analy- the root node and is the only functional hard maximum constraint ses (following refs. 3−5). Cordaixylon iowensis was not included in in BEAST runs using uniform priors, we carried out three runs the analyses because its assignment to the crown Acrogymno- with calibrations A through O (Table S4), and calibration P set to spermae already is supported by previous cladistic analyses (also [306.2, 351.7] (run 2), [306.2, 336.5] (run 3), and [306.2, 321.4] using the total-evidence approach) (6). Two data matrices were (run 4). The age estimates obtained in runs 2, 3, and 4 largely compiled. Matrix A comprised Ginkgo biloba, 12 living repre- overlapped with those from run 1 (Fig. S3). Second, we carried out two runs with different subsets of sentatives from each conifer family, and three fossils taxa related fi to Pinaceae and Araucariaceae (16 taxa in total; Fig. -

The Anatomy of the Bark of Libocedrus in New Zealand
IAWA Bulletin n.s., Vol. 6 (1),1985 23 THE ANATOMY OF THE BARK OF LlBOCEDRUS IN NEW ZEALAND by Lek-Lim Chan'" School of Forestry, University of Canterbury, Christchurch, New Zealand Summary The anatomy of the bark of Libocedrus bid southwards. In South Island it is also found in willii and L. plumosa, both indigenous to New lowland forests in the west. Libocedrus plumosa Zealand, is described. The phloem cell types in is also a tall tree and is found in lowland forest clude axial and ray parenchyma, sieve cells and from 35° to beyond 38° S in North Island and fibres. Fibres are of two types, thin-walled and between Collingwood and Westhaven (NW. Nel thick-walled. The ends of these fibres are most son) in South Island. ly blunt or abrupt. Minute crystals (crystal sand) were found in Terminology the radial walls of some axial parenchyma cells, 'Conducting phloem' refers to that part of sieve cells and fibres. Trabeculae were abun the phloem close to the vascular cambium where dant in the bark of L. bidwillii. sieve cells are still functioning in conduction, The walls of the phellem cells were very thin, and 'living bark' designates that part of the appearing flimsy. Phelloderm cells in L. plumo bark outside the vascular cambium up to and sa seem to undergo sclerification just prior to including the last-formed periderm. death, upon formation of a new periderm deep All other terms used for tissues of the bark er in the phloem. are in common usage (see IA WA, 1964; Society Key words: Bark anatomy, crystals, Libocedrus, of American Foresters, 1958). -
![NORFOLK RD NURSERY LTD PLANT LIST 2020 131 Norfolk Road,Carterton 5791 Phone: 06 370 2328 Mob: 027 634 7441 Email: Sales@Norfolknursery.Co.Nz Price [ GST Exclusive ]](https://docslib.b-cdn.net/cover/0771/norfolk-rd-nursery-ltd-plant-list-2020-131-norfolk-road-carterton-5791-phone-06-370-2328-mob-027-634-7441-email-sales-norfolknursery-co-nz-price-gst-exclusive-3470771.webp)
NORFOLK RD NURSERY LTD PLANT LIST 2020 131 Norfolk Road,Carterton 5791 Phone: 06 370 2328 Mob: 027 634 7441 Email: [email protected] Price [ GST Exclusive ]
NORFOLK RD NURSERY LTD PLANT LIST 2020 131 Norfolk Road,Carterton 5791 Phone: 06 370 2328 Mob: 027 634 7441 Email: [email protected] Price [ GST exclusive ] ** O/S meaning out of stock Botanical Name Common Name RTB .5L 1L 2.5L PB8 PB12 PB18 COMMERCIAL NATIVES - CAN BE ORDERED BY EMAIL OR OVER THE PHONE (SMALL GRADES) LOW-COST BULK QUANTITIES COMPETITIVE WHOLE SALE PRICE ECO SOURCED WAIRARAPA INDIGENOUS PLANTS SEE NON-COMMERCIAL LIST FOR LARGER GRADES Coprosma acerosa sand coprosma 3.21 7.39 O/S Coprosma areolata bush edge coprosma 7.39 Coprosma 'Autumn haze' 3.21 Coprosma crassifolia O/S Coprosma grandifolia kanono 3.21 7.39 Coprosma kirkii 3.21 7.39 Coprosma lucida shining karamu 5.65 O/S Coprsosma microcarpa O/S Coprosma pedicillata O/S Coprosma propinqua mingimingi 7.39 Coprosma Red Rocks 7.39 O/S Coprosma repens taupata 3.21 Coprosma rigida stiff karamu 7.39 Coprosma robusta karamu 2.6 Coprosma rugosa needle coprosma 3.21 7.39 O/S Coprosma tenuicaulis O/S Coprosma virescens 7,39 Cordyline australis cabbage tree; ti kouka 3.21 7.39 Corokia 'Blue Grey' 7.39 Corokia buddleioides korokio taranga 3.91 O/S Corokia cotoneaster korokio 7.39 Corokia macrocarpa hokotaka 7.39 Corokia x virgata hybrid corokia 7.39 Corokia ' Frosted Chocolate' 3.91 7.39 Corokia 'Genty's Green' 3.91 7.39 Corokia 'Grey Ghost' 7.39 Dacrycarpus dacrydioides kahikatea, white pine 3.21 7.39 Dodonaea viscosa akeake - Green 7.39 Dodonaea viscosa 'Purpurea' akeake - Red Griselinia littoralis Broadway mint 3.91 7.39 Griselinia 'Canterbury' 7.39 Griselinia littoralis -

1 Supporting Information Supplementary Methods the Main Purposes of Use of Wild Animal and Plant Species Recorded in the Red
1 Supporting information 2 3 Supplementary Methods 4 5 The main purposes of use of wild animal and plant species recorded in the Red List 6 7 We investigated the prevalence of different purposes of use from the use and trade information. 8 Because completing the Use and Trade classification scheme is not mandatory for Red List 9 assessors, we investigated the prevalence of Use and Trade coding to decide which species 10 groups to include in our analyses. We selected taxonomic groups for inclusion based on the 11 following criteria: i) >40% of all extant, data sufficient species, LC species and threatened 12 species have at least one purpose of use coded (thus selecting taxonomic groups with high 13 prevalence of use); and / or ii) the proportion of LC species with at least one purpose of use code 14 falls above or within the range of the proportion of species with Use and Trade coding across the 15 other Red List categories (thus also selecting taxonomic groups where use and trade may be 16 relatively low, but use and trade in LC species is coded to a similar level as that of species in 17 other RL categories). This limited our dataset to the following taxonomic groups which have 18 adequate recording of use and trade: birds, amphibians, reptiles, cycads, conifers and dicots from 19 the terrestrial group; and corals, bony fishes, crustaceans and cone snails from the aquatic species 20 group (Table S4). We excluded mammals, cephalopods and cartilaginous fishes as meeting 21 neither criteria i) nor ii), i.e. -

Bulletin of Thecupressus
BBuulllleettiinn of the CCuupprreessssuuss CCoonnsseerrvvaattiioonn PPrroojjeecctt No 10 Volume 4 No 3 ─ 19 December 2015 Contents Volume 4 Number 3 Morphology and morphogenesis of the seed cones of the Cupressaceae - part III: Callitroideae .......................................................................... 91 A. Jagel & V.M. Dörken Abstract : Morphology of the seed cones of the Callitroideae is given. These cones are significantly less variable than those of the Cupressoideae in respect to the number of cone scales, and especially the formation of the distal end of the cone axis. They consist only of two whorls of cone scales. In some taxa the cone axis is developed into a so called columella, which however does not reach as far as the apical surface of the cone, being shorter than the scales. In the Libocedrus group and in Diselma only the distal whorl is fertile. Ovules of the Callitroideae are always inserted axillary. The cone scale is formed only by the bract scale. Non-axillary ovules do not exist. In contrast to the Cupressoideae, in Actinostrobus and Callitris (and perhaps also in Fitzroya) ovules of the distal whorl are developed slightly earlier than those of the lower ones. Photo Gallery: Fitzroya cupressoides ..................................................................................... 109 J. Bisbee Photo Gallery: Libocedrus uvifera .......................................................................................... 116 J. Bisbee Photo Gallery: Austrocedrus chilensis ................................................................................... 124 J. Bisbee Note on the conservation status of Cupressus montana ........................................................ 133 D. Mаеrki Abstract : The conservation status of Cupressus montana a rare cypress species of the highest mountain range of Baja California is critically reviewed according to the available information. This Bulletin is edited by the Cupressus Conservation Project, a non-profit organisation based in Geneva, Switzerland. -

Conservation Status of New Zealand Indigenous Vascular Plants, 2012
NEW ZEALAND THREAT CLASSIFICATION SERIES 3 Conservation status of New Zealand indigenous vascular plants, 2012 Peter J. de Lange, Jeremy R. Rolfe, Paul D. Champion, Shannel P. Courtney, Peter B. Heenan, John W. Barkla, Ewen K. Cameron, David A. Norton and Rodney A. Hitchmough Cover: The Nationally Critical shrub Pittosporum serpentinum from the Surville Cliffs is severely affected by possums, and no seedlings have been found during recent surveys. Photo: Jeremy Rolfe. New Zealand Threat Classification Series is a scientific monograph series presenting publications related to the New Zealand Threat Classification System (NZTCS). Most will be lists providing NZTCS status of members of a plant or animal group (e.g. algae, birds, spiders). There are currently 23 groups, each assessed once every 3 years. After each 3-year cycle there will be a report analysing and summarising trends across all groups for that listing cycle. From time to time the manual that defines the categories, criteria and process for the NZTCS will be reviewed. Publications in this series are considered part of the formal international scientific literature. This report is available from the departmental website in pdf form. Titles are listed in our catalogue on the website, refer www.doc.govt.nz under Publications, then Science & technical. © Copyright August 2013, New Zealand Department of Conservation ISSN 2324–1713 (web PDF) ISBN 978–0–478–14995–1 (web PDF) This report was prepared for publication by the Publishing Team; editing by Amanda Todd and layout by Lynette Clelland. Publication was approved by the Deputy Director-General, Science and Capability Group, Department of Conservation, Wellington, New Zealand. -

Identification Key to the Cypress Family (Cupressaceae)1
Feddes Repertorium 116 (2005) 1–2, 96–146 DOI: 10.1002/fedr.200411062 Weinheim, Mai 2005 Ruhr-Universität Bochum, Lehrstuhl für Spezielle Botanik, Bochum C. SCHULZ; P. KNOPF & TH. STÜTZEL Identification key to the Cypress family (Cupressaceae)1 With 11 Figures Summary Zusammenfassung The identification of Cupressaceae taxa, except for Bestimmungsschlüssel für die Familie der Cup- some local and easily distinguishable taxa, is diffi- ressaceae cult even for specialists. One reason for this is the lack of a complete key including all Cupressaceae Die Bestimmung von Cupressaceae-Taxa ist mit taxa, another reason is that diagnoses and descrip- Ausnahme einiger lokaler und leicht bestimmbarer tions are spread over several hundred publications Taxa schwierig, selbst für Spezialisten. Ein Grund, which are sometimes difficult to access. Based on warum es noch keinen vollständigen Bestimmungs- morphological studies of about 3/4 of the species and schlüssel mit allen Cupressaceae-Taxa gibt ist, dass a careful compilation of the most important descrip- die Sippen-Beschreibungen sich auf mehrere hundert tions of Cupressaceae, a first identification key for Publikationen verteilen, welche teilweise schwierig the entire Cypress family (Cupressaceae) could be zu beschaffen sind. Etwa 3/4 der Cupressaceae-Ar- set up. The key comprises any of the 30 genera, 134 ten wurden morphologisch untersucht und die wich- species, 7 subspecies, 38 varieties, one form and thus tigsten Beschreibungen zusammengefasst, daraus all 180 taxa recognized by FARJON (2001). The key wurde dann der erste vollständige Bestimmungs- uses mainly features of adult leaves, female cones schlüssel für Cupressaceae erstellt. Der Bestim- and other characters which are all relatively easy to mungsschlüssel enthält 30 Gattungen, 134 Arten, be used. -

Plant Me Instead!
PLANT ME INSTEAD! WEST COAST NELSON/MARLBOROUGH Acknowledgements Thank you to the following people and organisations who helped with the production of this booklet: Lawrie Metcalf, Georgina Pahl (Mainly Natives Nursery), Mike Crawford (Nikau Gardens), Geoff Walls, Peter Russell, Peter Williams, Nick Ledgard; staff from Tasman District Council, Nelson City Council, Marlborough District Council, West Coast Regional Council, and Department of Conservation, for input, information and advice; John Barkla, Jeremy Rolfe, Trevor James, John Clayton, Peter de Lange, John Smith-Dodsworth, John Liddle (Liddle Wonder Nurseries), Geoff Bryant, Clayson Howell, John Sawyer and others who provided photos; Sonia Frimmel (What’s the Story) for design and layout. While all non-native alternatives have been screened against several databases to ensure they are not considered weedy, predicting future behaviour is not an exact science! The only way to be 100% sure is to use ecosourced native species. Published by: Weedbusters © 2007 ISBN: 978-0-9582844-1-7 Get rid of a weed, plant me instead! Many of the weedy species that are invading and damaging our natural areas are ornamental plants that have ‘jumped the fence’ from gardens and gone wild. It costs councils, government departments and private landowners millions of dollars, and volunteers and community groups thousands of unpaid hours, to control these weeds every year. This Plant Me Instead booklet profiles the environmental weeds of greatest concern to those in your region who work and volunteer in local parks and reserves, national parks, bush remnants, wetlands and coastal areas. Suggestions are given for locally-sold non-weedy species, both native and non- native, that can be used to replace these weeds in your garden. -

Southern Hemisphere Maps and Present-Day Nothofagus Distribution
Copyright is owned by the Author of the thesis. Permission is given for a copy to be downloaded by an individual for the purpose of research and private study only. The thesis may not be reproduced elsewhere without the permission of the Author. Discontinuous distributions of iconic New Zealand plant taxa and their implications for Southern Hemisphere biogeography A thesis presented in partial fulfilment of the requirements for the degree of Doctor of Philosophy in Plant Biology at Massey University, Palmerston North, New Zealand Michael Knapp January 2007 "With regard to general problems of biogeography, the biota of New Zealand has been, perhaps, the most important of any in the world. It has figured prominently in all discussions of austral biogeography, and all notable authorities have fe lt obliged to explain its history: explain New Zealand and the world fa lls into place aroundit. " Gareth Nelson (1975) Abstra ct Abstract New Zealand has long been regarded as a key to understanding discontinuous distributions in the Southern Hemisphere. The archipelago is a fragment of the ancient super continent Gondwana. It has been isolated for 80 million years, has an excellent fossil record, and some of its most ancient biota such as the Southern Beeches (Nothofagus) and the Araucariaceae show disjunct distribution patterns with relatives on other fragments of Gondwana. Some of the most controversial problems of Southern Hemisphere biogeography with wide ranging implications involve New Zealand taxa. Three of them have been addressed in this thesis. The transoceanic relationships of the genus Nothofagus have long been regarded as an iconic example of a distribution pattern resulting from the break up of Gondwana. -
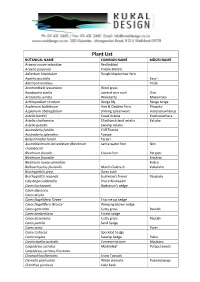
Plant List 2021
Plant List BOTANICAL NAME COMMON NAME MĀORI NAME Acaena novae-zelandiae Red bidibid Acaena purpurea Purple Bidibid Adiantum hispidulum Rough Maidenhair fern Agathis australis Kauri Alectryon excelsus Titoki Anemanthele lessoniana Wind grass Apodasmia similis Jointed wire rush Oioi Aristotelia serrata Wineberry Makomako Arthropodium cirratum Renga lily Renga renga Asplenium bulbiferum Hen & Chicken Fern Pikopiko Asplenium oblongifolium Shining spleenwort Huruhuruwhenua Astelia banksii Coast Astelia Kowharawhara Astelia chathamica Chatham Island astelia Kakaha Astelia grandis Swamp Astelia Austroderia fulvida Cliff Toetoe Austroderia splendens Toetoe Beilschmiedia tarairi Tarairi Austroblechnum lanceolatum (Blechnum Lance water fern Nini chambersii) Blechnum discolor Crown Fern Petipeti Blechnum fluviatile Kiwikiwi Blechnum novae-zelandiae Kiokio Bolboschoenus fluviatilis Marsh Clubrush Kukuraho Brachyglottis greyi Daisy bush Brachyglottis repanda bushman's friend Rangiora Calystegia soldanella Shore Bindweed Carex buchananii Buchanan's sedge Carex dipsacea Carex dissita Carex flagellifera 'Green' Trip me up sedge Carex flagellifera 'Bronze' Weeping brown sedge Carex geminata Cutty grass Rautahi Carex lambertiana Forest sedge Carex lessoniana Cutty grass Rautahi Carex pumila Sand Sedge Carex secta Purei Carex testacea Speckled Sedge Carex virgata Swamp Sedge Pukio Carmichaelia australis Common broom Maukoro Carpodetus serratus Marbleleaf Putaputaweta Carpodetus serratus Prostrata Chionochloa flavicans Snow Tussock Clematis paniculata White -

Threatened Plants of Waikato Conservancy
Teucridium parvifolium VERBENACEAE Status Gradual Decline Description A small-leaved shrub to 2 m. Young stems are orange or brown, square in cross-section and finely hairy. Leaves are in opposite pairs, round, dull green or brown-green; up to 12 mm long on stalks of equal length. Flowers are small (8 mm diameter), white (rarely bluish) with 5 irregular petal lobes. Fruit is a group of 4 seeds held within a persistent calyx. Flowering occurs from October to January; fruiting from December to March. Current record Historic record Extinct Similar species Small, round-leaved Coprosma species are similar, but easily distinguished by their round or oval stems, leaf domatia (or pits in the leaf near the midrib) and a small, pointed ‘stipule’ running across the stem between each leaf pair. Neomyrtus pedunculata has square stems, but they are hairless while the leaves are shiny and gland-dotted. Habitat Along stream sides and river terraces in lowland dry forest and podocarp-hardwood forest; occasionally on forest margins and amongst scrub. Distribution Endemic to New Zealand, occurring sporadically from Northland to Southland, but commoner in the east of both islands. In the Waikato, it occurs in several forest remnants around Waitomo. Threats Forest clearance, rural development, stock and feral animal browse, invasive weeds. Teucridium parvifolium. Photos: (above left) J.D. Sawyer; (left) S.P. Courtney. 78 Trilepidea adamsii Adams mistletoe LORANTHACEAE Status Extinct Description A shrubby, hemi-parasitic mistletoe up to 1 m in diameter. Parasitic on mamangi (Coprosma arborea), wharangi (Melicope ternata) and mapou (Myrsine australis). Leaves opposite, thick and fleshy, dark green, broadly elliptic or diamond-shaped, with paler green or reddish undersides. -

Structural Diversity and Function of Xyloglucan Sidechain Substituents
Supplementary Meterial Table S1. Xyloglucan branching patterns and sidechains of individual plant species. In the xylosylation motif column subscripts indicate the number of consecutive G units observed. Orders Family Species Generation/Organ/Tissue Xylosylation Motif Sidechains Reference Liverworts Marchantiales Marchantiaceae Marchantia polymorpha Gametophyte XXGG X, L, P, Q [1] Mosses Funariales Funariaceae Physcomitrella patens Protonema, gametophore XXGG3 X, L, M, N, P, Q [1] Hornworts Dendrocerotales Dendrocerotaceae Megaceros sp. Gametophyte XXXG X, L, F [1] Notothyladales Notothyladaceae Phaeoceros sp. Gametophyte XXXG X, L, F [1] Anthocerotales Anthocerotaceae Anthoceros agrestis Gametophyte XXXG X, L, F [1] Lycophytes Lycopodiales Huperziaceae Huperzia lucidula Sporophyte XXXG, XXGG X, L, D [1] Lycopodiaceae Lycopodium tristachyum Sporophyte XXXG, XXGG X, L, F, D, E [1] Lycopodiaceae Lycopodium cernuum Aerial stems with leaves XXGG2–5, XXXG X, L [2] Selaginellales Selaginellaceae Selaginella kraussiana Sporophyte XXXG X, L, F, D, E [1] Stems with leaves XXXG, X, L, F, D, E [2] Ferns Equisetales Equisetaceae Equisetum hyemale Sporophyte XXXG X, L, F, D, E, S [1] Stem internodes XXXG X, L, D, E, [2] Psilotales Psilotaceae Psilotum nudum Sporophyte XXXG X, L, F [1] Upper forked aerial stems XXGG2–3 X, L, F [2] Polypodiales Polypodiaceae Platycerium bifurcatum Sporophyte XXXG X, L, F [1] Pteridaceae Ceratopteris richardii Sporophyte XXXG X, L, F, S [1] Marattiales Marattiaceae Marattia salicina Fertile pinnae XXXG X, L, F [2] Osmundales Osmundaceae Osmunda regalis Lower sterile pinnae XXXG X, L, F [2] Hymenophyllales Hymenophyllaceae Trichomanes reniforme Laminae XXXG X, L, F, M [2] Hymenophyllaceae Hymenophyllum demissum Laminae XXXG X, L, F [2] Plants 2014, 3 S2 Table S1.