Hypotheses for the Decline of Cod in the North Atlantic*
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Characterization of Effluent from an Inland, Low- Salinity Shrimp Farm
Aquacultural Engineering 27 (2003) 147Á/156 www.elsevier.com/locate/aqua-online Characterization of effluent from an inland, low- salinity shrimp farm: what contribution could this water make if used for irrigation Dennis McIntosh ,Kevin Fitzsimmons Environmental Research Lab, University of Arizona, 2601 East Airport Drive, Tucson, AZ 85706, USA Received 7 July 2002; accepted 7 October 2002 Abstract Coastal aquaculture can contribute to eutrophication of receiving waters. New technologies and improved management practices allow the aquaculture industry to be more sustainable and economically viable. Current practices, however, do not provide an additional use for effluent water. Nitrogen, phosphorus and other effluent compounds could be valuable plant nutrients. Inflow and effluent water from an inland, low-salinity shrimp farm, were monitored. Bi-weekly analysis included total nitrogen, ammonia-nitrogen, nitrite-nitrogen, nitrate- nitrogen, total phosphorus, reactive phosphorus, alkalinity, chemical oxygen demand (COD), biochemical oxygen demand (BOD), total suspended solids (TSS) and volatile suspended solids (VSS), as well as temperature, salinity, dissolved oxygen and pH. Alkalinity and total nitrogen decreased during the in-pond residency. The other parameters increased while in the ponds. The potential benefit of having nutrient enriched wastewater to irrigate field crops was substantial, supplying between 20 and 31% of the necessary nitrogen fertilizer for wheat production. # 2002 Elsevier Science B.V. All rights reserved. Keywords: Inland aquaculture; Integration; Arizona; Marine shrimp Corresponding author. Tel.: /1-520-626-3318; fax: /1-520-573-0852 E-mail address: [email protected] (D. McIntosh). 0144-8609/02/$ - see front matter # 2002 Elsevier Science B.V. All rights reserved. PII: S 0 1 4 4 - 8 6 0 9 ( 0 2 ) 0 0 0 5 4 - 7 148 D. -

Cod Fact Sheet
R FACT SHEET Cod (Gadus morhua) Image taken from (Cohen et al. 1990) Introduction Cod (Gadus morhua) is generally considered a demersal fish although its habitat may become pelagic under certain hydrographic conditions, when feeding or spawning. It is widely distributed throughout the north Atlantic and Arctic regions in a variety of habitats from shoreline to continental shelf, in depths to 600m (Cohen et al. 1990). The Irish Sea stock spawns at two main sites in the western and eastern Irish Sea during February to April (Armstrong et al. 2011). Historically the stock has been commercially important, however in the last decade a decline in SSB and reduced productivity of the stock have led to reduce landings. Reviewed distribution map for Gadus morhua (Atlantic cod), with modelled year 2100 native range map based on IPCC A2 emissions scenario. www.aquamaps.org, version of Aug. 2013. Web. Accessed 28 Jan. 2011 Life history overview Adults are usually found in deeper, colder waters. During the day they form schools and swim about 30-80 m above the bottom, dispersing at night to feed (Cohen et al. 1990; ICES 2005). They are omnivorous; feeding at dawn or dusk on invertebrates and fish, including their own young (Cohen et al. 1990). Adults migrate between spawning, feeding and overwintering areas, mostly within the boundaries of the respective stocks. Large migrations are rare occurrences, although there is evidence for limited seasonal migrations into neighbouring regions, most Irish Sea fish will stay within their management area (ICES 2012). Historical tagging studies indicated spawning site fidelity but with varying degrees of mixing of cod between the Irish Sea, Celtic Sea and west of Scotland/north of Ireland (ICES 2015). -

Do Some Atlantic Bluefin Tuna Skip Spawning?
SCRS/2006/088 Col. Vol. Sci. Pap. ICCAT, 60(4): 1141-1153 (2007) DO SOME ATLANTIC BLUEFIN TUNA SKIP SPAWNING? David H. Secor1 SUMMARY During the spawning season for Atlantic bluefin tuna, some adults occur outside known spawning centers, suggesting either unknown spawning regions, or fundamental errors in our current understanding of bluefin tuna reproductive schedules. Based upon recent scientific perspectives, skipped spawning (delayed maturation and non-annual spawning) is possibly prevalent in moderately long-lived marine species like bluefin tuna. In principle, skipped spawning represents a trade-off between current and future reproduction. By foregoing reproduction, an individual can incur survival and growth benefits that accrue in deferred reproduction. Across a range of species, skipped reproduction was positively correlated with longevity, but for non-sturgeon species, adults spawned at intervals at least once every two years. A range of types of skipped spawning (constant, younger, older, event skipping; and delays in first maturation) was modeled for the western Atlantic bluefin tuna population to test for their effects on the egg-production-per-recruit biological reference point (stipulated at 20% and 40%). With the exception of extreme delays in maturation, skipped spawning had relatively small effect in depressing fishing mortality (F) threshold values. This was particularly true in comparison to scenarios of a juvenile fishery (ages 4-7), which substantially depressed threshold F values. Indeed, recent F estimates for 1990-2002 western Atlantic bluefin tuna stock assessments were in excess of threshold F values when juvenile size classes were exploited. If western bluefin tuna are currently maturing at an older age than is currently assessed (i.e., 10 v. -

Our Nation's Fisheries Will Be a Lot Easier Once We've Used up Everything Except Jellyfish!
Mismanaging Our Nation’s Fisheriesa menu of what's missing Limited quantity: get ‘em while supplies last Ted Stevens Alaskan Surprise Due to years of overfishing, we probably won’t be serving up Pacific Ocean perch, Tanner crab, Greenland turbot or rougheye rockfish. They may be a little hard to swallow, but Senator Stevens and the North Pacific Council will be sure to offer last minute riders, father and son sweetheart deals, record-breaking quotas, industry-led research, conflicts of interest and anti-trust violations. Meanwhile, fur seals, sea lions and sea otters are going hungry and disappearing fast. Surprise! Pacific Rockfish: See No Fish, Eat No Fish Cowcod, Canary Rockfish and Bocaccio are just three examples of rockfish managed by the Pacific Council that are overfished. As for the exact number of West Coast groundfish that are overfished, who knows? Without surveys to tell them what’s going on, what are they managing exactly? Striped Bass: Thin Is In! This popular Atlantic rockfish is available in abundance. Unfortunately, many appear to be undernourished and suffering from lesions – a condition that may point to Omega Protein’s industrial fishery of menhaden, the striper’s favorite prey species. Actually, you may want to hold off on this one until ASMFC starts regulating menhaden. Can you believe there are still no catch limits? Red Snapper Bycatch Platter While we are unable to provide full-size red snapper, we offer this plate of twenty juvenile red snapper discarded as bycatch from a Gulf of Mexico shrimp trawler for your dining pleasure. Shrimpers take and throw away about half of all young red snappers along the Texas coast, so we’ll keep these little guys coming straight from the back of the boat to the back of your throat! Caribbean Reef Fish Grab Bag What’s for dinner from the Caribbean? Who knows? With coral reefs in their jurisdiction, you would expect the Caribbean Council to be pioneering the ecosystem-based management approach and implementing the precautionary principle approach. -

Probable Discards of Cod in the Barents Sea and Ajacent Waters During Russian Bottom Trawl
unuftp.is Final Project 2002 PROBABLE DISCARDS OF COD IN THE BARENTS SEA AND AJACENT WATERS DURING RUSSIAN BOTTOM TRAWL Konstantin Sokolov Knipovich Polar Research Institute of Marine Fisheries and Oceanography (PINRO) Murmansk, Russia [email protected] Supervisor Dr. Palsson, O.K. Marine Research Institute Reykjavik, Iceland ABSTRACT Unaccounted discarding of small-sized fish is an important and acute problem in many fisheries because it affects the condition of stocks and reduces the reliability of fishery statistics. Such discards are regarded as a threat to intensively exploited species, such as the North-East Arctic cod (Gadus morhua morhua L.) which inhabits the Barents Sea and adjacent waters. In the present study an attempt is made to estimate of the quantity of small-sized cod discarded in the Russian bottom trawl fishery in 1996-2001. This work is based on cod length measurements onboard Russian commercial vessels in the period 1996-2001 and distributional features, such as density and size composition inferred from catch statistics. The calculated annual discards of small cod were estimated to be in the range of 3-80 million individuals. Discards appeared to be highest in 1998 and lowest in 1996 and 2001. This was found to be related to the abundance of cod recruits and the portion of total catch taken in the Eastern-Central part of the Barents Sea. The features of spatial and seasonal distribution of small-sized cod, the depth and duration of trawling and catch of cod influence the catch of small cod per unit of effort and, as a consequence, discards of such fish. -

The Landing Obligation and Its Implications on the Control of Fisheries
DIRECTORATE-GENERAL FOR INTERNAL POLICIES POLICY DEPARTMENT B: STRUCTURAL AND COHESION POLICIES FISHERIES THE LANDING OBLIGATION AND ITS IMPLICATIONS ON THE CONTROL OF FISHERIES STUDY This document was requested by the European Parliament's Committee on Fisheries. AUTHORS Ocean Governance Consulting: Christopher Hedley Centre for Environment, Fisheries and Aquaculture Science: Tom Catchpole, Ana Ribeiro Santos RESPONSIBLE ADMINISTRATOR Marcus Breuer Policy Department B: Structural and Cohesion Policies European Parliament B-1047 Brussels E-mail: [email protected] EDITORIAL ASSISTANCE Adrienn Borka Lyna Pärt LINGUISTIC VERSIONS Original: EN ABOUT THE PUBLISHER To contact the Policy Department or to subscribe to its monthly newsletter please write to: [email protected] Manuscript completed in September 2015. © European Union, 2015. Print ISBN 978-92-823-7938-7 doi:10.2861/694624 QA-02-15-709-EN-C PDF ISBN 978-92-823-7939-4 doi:10.2861/303902 QA-02-15-709-EN-N This document is available on the Internet at: http://www.europarl.europa.eu/studies DISCLAIMER The opinions expressed in this document are the sole responsibility of the author and do not necessarily represent the official position of the European Parliament. Reproduction and translation for non-commercial purposes are authorized, provided the source is acknowledged and the publisher is given prior notice and sent a copy. DIRECTORATE-GENERAL FOR INTERNAL POLICIES POLICY DEPARTMENT B: STRUCTURAL AND COHESION POLICIES FISHERIES THE LANDING OBLIGATION AND ITS IMPLICATIONS ON THE CONTROL OF FISHERIES STUDY Abstract This study reviews the impacts of the new Common Fisheries Policy (CFP) rules requiring catches in regulated fisheries to be landed and counted against quotas of each Member State ("the landing obligation and requiring that catch of species subject to the landing obligation below a minimum conservation reference size be restricted to purposes other than direct human consumption. -

Wholesale Market Profiles for Alaska Groundfish and Crab Fisheries
JANUARY 2020 Wholesale Market Profiles for Alaska Groundfish and FisheriesCrab Wholesale Market Profiles for Alaska Groundfish and Crab Fisheries JANUARY 2020 JANUARY Prepared by: McDowell Group Authors and Contributions: From NOAA-NMFS’ Alaska Fisheries Science Center: Ben Fissel (PI, project oversight, project design, and editor), Brian Garber-Yonts (editor). From McDowell Group, Inc.: Jim Calvin (project oversight and editor), Dan Lesh (lead author/ analyst), Garrett Evridge (author/analyst) , Joe Jacobson (author/analyst), Paul Strickler (author/analyst). From Pacific States Marine Fisheries Commission: Bob Ryznar (project oversight and sub-contractor management), Jean Lee (data compilation and analysis) This report was produced and funded by the NOAA-NMFS’ Alaska Fisheries Science Center. Funding was awarded through a competitive contract to the Pacific States Marine Fisheries Commission and McDowell Group, Inc. The analysis was conducted during the winter of 2018 and spring of 2019, based primarily on 2017 harvest and market data. A final review by staff from NOAA-NMFS’ Alaska Fisheries Science Center was completed in June 2019 and the document was finalized in March 2016. Data throughout the report was compiled in November 2018. Revisions to source data after this time may not be reflect in this report. Typically, revisions to economic fisheries data are not substantial and data presented here accurately reflects the trends in the analyzed markets. For data sourced from NMFS and AKFIN the reader should refer to the Economic Status Report of the Groundfish Fisheries Off Alaska, 2017 (https://www.fisheries.noaa.gov/resource/data/2017-economic-status-groundfish-fisheries-alaska) and Economic Status Report of the BSAI King and Tanner Crab Fisheries Off Alaska, 2018 (https://www.fisheries.noaa. -
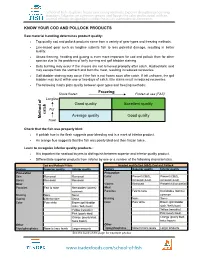
Know Your Cod and Pollock Products
School of Fish - Explores frozen processing methods, Exposes deceptive processing practices, Educates on common species and Equips the sales professional with an arsenal of tools designed to combat the issues vulnerable to deception. KNOW YOUR COD AND POLLOCK PRODUCTS Raw material handling determines product quality: • Top quality cod and pollock products come from a variety of gear types and freezing methods. • Line-based gear such as longline submits fish to less potential damage, resulting in better quality. • At-sea freezing, heading and gutting is even more important for cod and pollock than for other species due to the problems of belly burning and gall bladder staining. • Belly burning may occur if the viscera are not removed promptly after catch. Hydrochloric acid may escape from the stomach and burn the meat, resulting in reduced recoveries. • Gall bladder staining may occur if the fish is not frozen soon after catch. If left unfrozen, the gall bladder may burst within one or two days of catch. Bile stains result in reduced recoveries. • The following matrix plots quality between gear types and freezing methods: Shore frozen Freezing Frozen at sea (FAS) Longline of Good quality Excellent quality Pot catch Average quality Good quality Method Trawl Check that the fish was properly bled: • A pinkish hue in the flesh suggests poor bleeding and is a mark of inferior product. • An orange hue suggests that the fish was poorly bled and then frozen twice. Learn to recognize inferior quality products: • It is important for seafood buyers to distinguish between superior and inferior quality product. • Differentiate superior products from inferior by one or a number of the following characteristics. -

The Decline of Atlantic Cod – a Case Study
The Decline of Atlantic Cod – A Case Study Author contact information Wynn W. Cudmore, Ph.D., Principal Investigator Northwest Center for Sustainable Resources Chemeketa Community College P.O. Box 14007 Salem, OR 97309 E-mail: [email protected] Phone: 503-399-6514 Published 2009 DUE # 0757239 1 NCSR curriculum modules are designed as comprehensive instructions for students and supporting materials for faculty. The student instructions are designed to facilitate adaptation in a variety of settings. In addition to the instructional materials for students, the modules contain separate supporting information in the "Notes to Instructors" section, and when appropriate, PowerPoint slides. The modules also contain other sections which contain additional supporting information such as assessment strategies and suggested resources. The PowerPoint slides associated with this module are the property of the Northwest Center for Sustainable Resources (NCSR). Those containing text may be reproduced and used for any educational purpose. Slides with images may be reproduced and used without prior approval of NCSR only for educational purposes associated with this module. Prior approval must be obtained from NCSR for any other use of these images. Permission requests should be made to [email protected]. Acknowledgements We thank Bill Hastie of Northwest Aquatic and Marine Educators (NAME), and Richard O’Hara of Chemeketa Community College for their thoughtful reviews. Their comments and suggestions greatly improved the quality of this module. We thank NCSR administrative assistant, Liz Traver, for the review, graphic design and layout of this module. 2 Table of Contents NCSR Marine Fisheries Series ....................................................................................................... 4 The Decline of Atlantic Cod – A Case Study ................................................................................ -

Atlantic Cod (Gadus Morhua) Off Newfoundland and Labrador Determined from Genetic Variation
COSEWIC Assessment and Update Status Report on the Atlantic Cod Gadus morhua Newfoundland and Labrador population Laurentian North population Maritimes population Arctic population in Canada Newfoundland and Labrador population - Endangered Laurentian North population - Threatened Maritimes population - Special Concern Arctic population - Special Concern 2003 COSEWIC COSEPAC COMMITTEE ON THE STATUS OF COMITÉ SUR LA SITUATION ENDANGERED WILDLIFE DES ESPÈCES EN PÉRIL IN CANADA AU CANADA COSEWIC status reports are working documents used in assigning the status of wildlife species suspected of being at risk. This report may be cited as follows: COSEWIC 2003. COSEWIC assessment and update status report on the Atlantic cod Gadus morhua in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. xi + 76 pp. Production note: COSEWIC would like to acknowledge Jeffrey A. Hutchings for writing the update status report on the Atlantic cod Gadus morhua, prepared under contract with Environment Canada. For additional copies contact: COSEWIC Secretariat c/o Canadian Wildlife Service Environment Canada Ottawa, ON K1A 0H3 Tel.: (819) 997-4991 / (819) 953-3215 Fax: (819) 994-3684 E-mail: COSEWIC/[email protected] http://www.cosewic.gc.ca Également disponible en français sous le titre Rapport du COSEPAC sur la situation de la morue franche (Gadus morhua) au Canada Cover illustration: Atlantic Cod — Line drawing of Atlantic cod Gadus morhua by H.L. Todd. Image reproduced with permission from the Smithsonian Institution, NMNH, Division of Fishes. Her Majesty the Queen in Right of Canada, 2003 Catalogue No.CW69-14/311-2003-IN ISBN 0-662-34309-3 Recycled paper COSEWIC Assessment Summary Assessment summary — May 2003 Common name Atlantic cod (Newfoundland and Labrador population) Scientific name Gadus morhua Status Endangered Reason for designation Cod in the inshore and offshore waters of Labrador and northeastern Newfoundland, including Grand Bank, having declined 97% since the early 1970s and more than 99% since the early 1960s, are now at historically low levels. -

The Commonwealth of Massachusetts Division of Marine Fisheries
The Commonwealth of Massachusetts Division of Marine Fisheries 251 Causeway Street, Suite 400, Boston, MA 02114 p: (617) 626-1520 | f: (617) 626-1509 www.mass.gov/marinefisheries CHARLES D. BAKER KARYN E. POLITO KATHLEEN A. THEOHARIDES RONALD S. AMIDON DANIEL J. MCKIERNAN Governor Lt. Governor Secretary Commissioner Director February 19, 2021 MarineFisheries Advisory New Protected Species Regulations Finalized for Fixed Gear Fisheries and Industry Outreach on Required Gear Modifications This advisory serves to provide you with information regarding new protected species regulations. This includes a description of the new regulatory requirements, resources available to assist commercial trap fishermen in complying with the new gear modification and weak rope rules, and additional information germane to DMF’s ongoing efforts to protect the North Atlantic right whale. New Regulations Enacted The Marine Fisheries Advisory Commission approved several new regulatory measures at their January 28, 2021 business meeting affecting protected species and fixed gear fishing in Massachusetts (Advisory). These regulations have been filed with the Secretary of State and will be implemented as of March 5, 2021. The seasonal closure rules will be effective March 5, 2021, whereas the gear modification rules will not go into effect until May 2021 to provide fishermen with time to alter their gear configuration. More details on each new regulatory provision are provided in the bullets below. • Seasonal Commercial Trap Gear Closure. A new commercial trap gear closure goes into effect on March 5, 2021. This new regulation extends the prior seasonal state waters commercial trap gear closure in both space and time. The prior closure occurred from February 1 – April 30 within Cape Cod Bay, Stellwagen Bank, and the Outer Cape Cod Lobster Management Area. -

Abundance and Distribution of Atlantic Cod (Gadus Morhua)
145 National Marine Fisheries Service Fishery Bulletin First U.S. Commissioner established in 1881 of Fisheries and founder NOAA of Fishery Bulletin Abstract—Atlantic cod (Gadus morhua) Abundance and distribution of Atlantic cod in southern New England (SNE) and along the mid-Atlantic coast have been (Gadus morhua) in a warming southern described as the world’s southernmost population of this species, but little New England is known of their population dynam- ics. Despite the expectation that SNE Joseph A. Langan (contact author)1 Atlantic cod are or will be negatively M. Conor McManus2 influenced by increasing water tem- 3 peratures due to climate change, fish- Douglas R. Zemeckis 1 eries that target Atlantic cod in this Jeremy S. Collie region have reported increased land- ings during the past 2 decades. The Email address for contact author: [email protected] work described here used ichthyoplank- ton and trawl survey data to investigate 1 Graduate School of Oceanography 3 Department of Agriculture and Natural spatial and temporal patterns of abun- University of Rhode Island Resources dance of Atlantic cod, and their potential Narragansett Bay Campus New Jersey Agricultural Experiment Station links to environmental factors, across 215 South Ferry Road Rutgers, the State University of New Jersey multiple life stages in Rhode Island. Narragansett, Rhode Island 02882 1623 Whitesville Road The results identify waters of the state Toms River, New Jersey 08755 of Rhode Island as a settlement and 2 Division of Marine Fisheries nursery area for early stages of Atlantic Rhode Island Department of Environmental cod until water temperatures approach Management 15°C in late spring.