Abundance and Distribution of Atlantic Cod (Gadus Morhua)
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Characterization of Effluent from an Inland, Low- Salinity Shrimp Farm
Aquacultural Engineering 27 (2003) 147Á/156 www.elsevier.com/locate/aqua-online Characterization of effluent from an inland, low- salinity shrimp farm: what contribution could this water make if used for irrigation Dennis McIntosh ,Kevin Fitzsimmons Environmental Research Lab, University of Arizona, 2601 East Airport Drive, Tucson, AZ 85706, USA Received 7 July 2002; accepted 7 October 2002 Abstract Coastal aquaculture can contribute to eutrophication of receiving waters. New technologies and improved management practices allow the aquaculture industry to be more sustainable and economically viable. Current practices, however, do not provide an additional use for effluent water. Nitrogen, phosphorus and other effluent compounds could be valuable plant nutrients. Inflow and effluent water from an inland, low-salinity shrimp farm, were monitored. Bi-weekly analysis included total nitrogen, ammonia-nitrogen, nitrite-nitrogen, nitrate- nitrogen, total phosphorus, reactive phosphorus, alkalinity, chemical oxygen demand (COD), biochemical oxygen demand (BOD), total suspended solids (TSS) and volatile suspended solids (VSS), as well as temperature, salinity, dissolved oxygen and pH. Alkalinity and total nitrogen decreased during the in-pond residency. The other parameters increased while in the ponds. The potential benefit of having nutrient enriched wastewater to irrigate field crops was substantial, supplying between 20 and 31% of the necessary nitrogen fertilizer for wheat production. # 2002 Elsevier Science B.V. All rights reserved. Keywords: Inland aquaculture; Integration; Arizona; Marine shrimp Corresponding author. Tel.: /1-520-626-3318; fax: /1-520-573-0852 E-mail address: [email protected] (D. McIntosh). 0144-8609/02/$ - see front matter # 2002 Elsevier Science B.V. All rights reserved. PII: S 0 1 4 4 - 8 6 0 9 ( 0 2 ) 0 0 0 5 4 - 7 148 D. -

Cod Fact Sheet
R FACT SHEET Cod (Gadus morhua) Image taken from (Cohen et al. 1990) Introduction Cod (Gadus morhua) is generally considered a demersal fish although its habitat may become pelagic under certain hydrographic conditions, when feeding or spawning. It is widely distributed throughout the north Atlantic and Arctic regions in a variety of habitats from shoreline to continental shelf, in depths to 600m (Cohen et al. 1990). The Irish Sea stock spawns at two main sites in the western and eastern Irish Sea during February to April (Armstrong et al. 2011). Historically the stock has been commercially important, however in the last decade a decline in SSB and reduced productivity of the stock have led to reduce landings. Reviewed distribution map for Gadus morhua (Atlantic cod), with modelled year 2100 native range map based on IPCC A2 emissions scenario. www.aquamaps.org, version of Aug. 2013. Web. Accessed 28 Jan. 2011 Life history overview Adults are usually found in deeper, colder waters. During the day they form schools and swim about 30-80 m above the bottom, dispersing at night to feed (Cohen et al. 1990; ICES 2005). They are omnivorous; feeding at dawn or dusk on invertebrates and fish, including their own young (Cohen et al. 1990). Adults migrate between spawning, feeding and overwintering areas, mostly within the boundaries of the respective stocks. Large migrations are rare occurrences, although there is evidence for limited seasonal migrations into neighbouring regions, most Irish Sea fish will stay within their management area (ICES 2012). Historical tagging studies indicated spawning site fidelity but with varying degrees of mixing of cod between the Irish Sea, Celtic Sea and west of Scotland/north of Ireland (ICES 2015). -

Do Some Atlantic Bluefin Tuna Skip Spawning?
SCRS/2006/088 Col. Vol. Sci. Pap. ICCAT, 60(4): 1141-1153 (2007) DO SOME ATLANTIC BLUEFIN TUNA SKIP SPAWNING? David H. Secor1 SUMMARY During the spawning season for Atlantic bluefin tuna, some adults occur outside known spawning centers, suggesting either unknown spawning regions, or fundamental errors in our current understanding of bluefin tuna reproductive schedules. Based upon recent scientific perspectives, skipped spawning (delayed maturation and non-annual spawning) is possibly prevalent in moderately long-lived marine species like bluefin tuna. In principle, skipped spawning represents a trade-off between current and future reproduction. By foregoing reproduction, an individual can incur survival and growth benefits that accrue in deferred reproduction. Across a range of species, skipped reproduction was positively correlated with longevity, but for non-sturgeon species, adults spawned at intervals at least once every two years. A range of types of skipped spawning (constant, younger, older, event skipping; and delays in first maturation) was modeled for the western Atlantic bluefin tuna population to test for their effects on the egg-production-per-recruit biological reference point (stipulated at 20% and 40%). With the exception of extreme delays in maturation, skipped spawning had relatively small effect in depressing fishing mortality (F) threshold values. This was particularly true in comparison to scenarios of a juvenile fishery (ages 4-7), which substantially depressed threshold F values. Indeed, recent F estimates for 1990-2002 western Atlantic bluefin tuna stock assessments were in excess of threshold F values when juvenile size classes were exploited. If western bluefin tuna are currently maturing at an older age than is currently assessed (i.e., 10 v. -

Our Nation's Fisheries Will Be a Lot Easier Once We've Used up Everything Except Jellyfish!
Mismanaging Our Nation’s Fisheriesa menu of what's missing Limited quantity: get ‘em while supplies last Ted Stevens Alaskan Surprise Due to years of overfishing, we probably won’t be serving up Pacific Ocean perch, Tanner crab, Greenland turbot or rougheye rockfish. They may be a little hard to swallow, but Senator Stevens and the North Pacific Council will be sure to offer last minute riders, father and son sweetheart deals, record-breaking quotas, industry-led research, conflicts of interest and anti-trust violations. Meanwhile, fur seals, sea lions and sea otters are going hungry and disappearing fast. Surprise! Pacific Rockfish: See No Fish, Eat No Fish Cowcod, Canary Rockfish and Bocaccio are just three examples of rockfish managed by the Pacific Council that are overfished. As for the exact number of West Coast groundfish that are overfished, who knows? Without surveys to tell them what’s going on, what are they managing exactly? Striped Bass: Thin Is In! This popular Atlantic rockfish is available in abundance. Unfortunately, many appear to be undernourished and suffering from lesions – a condition that may point to Omega Protein’s industrial fishery of menhaden, the striper’s favorite prey species. Actually, you may want to hold off on this one until ASMFC starts regulating menhaden. Can you believe there are still no catch limits? Red Snapper Bycatch Platter While we are unable to provide full-size red snapper, we offer this plate of twenty juvenile red snapper discarded as bycatch from a Gulf of Mexico shrimp trawler for your dining pleasure. Shrimpers take and throw away about half of all young red snappers along the Texas coast, so we’ll keep these little guys coming straight from the back of the boat to the back of your throat! Caribbean Reef Fish Grab Bag What’s for dinner from the Caribbean? Who knows? With coral reefs in their jurisdiction, you would expect the Caribbean Council to be pioneering the ecosystem-based management approach and implementing the precautionary principle approach. -
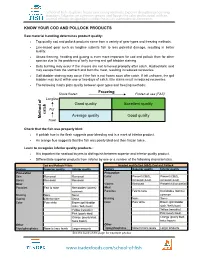
Know Your Cod and Pollock Products
School of Fish - Explores frozen processing methods, Exposes deceptive processing practices, Educates on common species and Equips the sales professional with an arsenal of tools designed to combat the issues vulnerable to deception. KNOW YOUR COD AND POLLOCK PRODUCTS Raw material handling determines product quality: • Top quality cod and pollock products come from a variety of gear types and freezing methods. • Line-based gear such as longline submits fish to less potential damage, resulting in better quality. • At-sea freezing, heading and gutting is even more important for cod and pollock than for other species due to the problems of belly burning and gall bladder staining. • Belly burning may occur if the viscera are not removed promptly after catch. Hydrochloric acid may escape from the stomach and burn the meat, resulting in reduced recoveries. • Gall bladder staining may occur if the fish is not frozen soon after catch. If left unfrozen, the gall bladder may burst within one or two days of catch. Bile stains result in reduced recoveries. • The following matrix plots quality between gear types and freezing methods: Shore frozen Freezing Frozen at sea (FAS) Longline of Good quality Excellent quality Pot catch Average quality Good quality Method Trawl Check that the fish was properly bled: • A pinkish hue in the flesh suggests poor bleeding and is a mark of inferior product. • An orange hue suggests that the fish was poorly bled and then frozen twice. Learn to recognize inferior quality products: • It is important for seafood buyers to distinguish between superior and inferior quality product. • Differentiate superior products from inferior by one or a number of the following characteristics. -

The Decline of Atlantic Cod – a Case Study
The Decline of Atlantic Cod – A Case Study Author contact information Wynn W. Cudmore, Ph.D., Principal Investigator Northwest Center for Sustainable Resources Chemeketa Community College P.O. Box 14007 Salem, OR 97309 E-mail: [email protected] Phone: 503-399-6514 Published 2009 DUE # 0757239 1 NCSR curriculum modules are designed as comprehensive instructions for students and supporting materials for faculty. The student instructions are designed to facilitate adaptation in a variety of settings. In addition to the instructional materials for students, the modules contain separate supporting information in the "Notes to Instructors" section, and when appropriate, PowerPoint slides. The modules also contain other sections which contain additional supporting information such as assessment strategies and suggested resources. The PowerPoint slides associated with this module are the property of the Northwest Center for Sustainable Resources (NCSR). Those containing text may be reproduced and used for any educational purpose. Slides with images may be reproduced and used without prior approval of NCSR only for educational purposes associated with this module. Prior approval must be obtained from NCSR for any other use of these images. Permission requests should be made to [email protected]. Acknowledgements We thank Bill Hastie of Northwest Aquatic and Marine Educators (NAME), and Richard O’Hara of Chemeketa Community College for their thoughtful reviews. Their comments and suggestions greatly improved the quality of this module. We thank NCSR administrative assistant, Liz Traver, for the review, graphic design and layout of this module. 2 Table of Contents NCSR Marine Fisheries Series ....................................................................................................... 4 The Decline of Atlantic Cod – A Case Study ................................................................................ -

Atlantic Cod (Gadus Morhua) Off Newfoundland and Labrador Determined from Genetic Variation
COSEWIC Assessment and Update Status Report on the Atlantic Cod Gadus morhua Newfoundland and Labrador population Laurentian North population Maritimes population Arctic population in Canada Newfoundland and Labrador population - Endangered Laurentian North population - Threatened Maritimes population - Special Concern Arctic population - Special Concern 2003 COSEWIC COSEPAC COMMITTEE ON THE STATUS OF COMITÉ SUR LA SITUATION ENDANGERED WILDLIFE DES ESPÈCES EN PÉRIL IN CANADA AU CANADA COSEWIC status reports are working documents used in assigning the status of wildlife species suspected of being at risk. This report may be cited as follows: COSEWIC 2003. COSEWIC assessment and update status report on the Atlantic cod Gadus morhua in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. xi + 76 pp. Production note: COSEWIC would like to acknowledge Jeffrey A. Hutchings for writing the update status report on the Atlantic cod Gadus morhua, prepared under contract with Environment Canada. For additional copies contact: COSEWIC Secretariat c/o Canadian Wildlife Service Environment Canada Ottawa, ON K1A 0H3 Tel.: (819) 997-4991 / (819) 953-3215 Fax: (819) 994-3684 E-mail: COSEWIC/[email protected] http://www.cosewic.gc.ca Également disponible en français sous le titre Rapport du COSEPAC sur la situation de la morue franche (Gadus morhua) au Canada Cover illustration: Atlantic Cod — Line drawing of Atlantic cod Gadus morhua by H.L. Todd. Image reproduced with permission from the Smithsonian Institution, NMNH, Division of Fishes. Her Majesty the Queen in Right of Canada, 2003 Catalogue No.CW69-14/311-2003-IN ISBN 0-662-34309-3 Recycled paper COSEWIC Assessment Summary Assessment summary — May 2003 Common name Atlantic cod (Newfoundland and Labrador population) Scientific name Gadus morhua Status Endangered Reason for designation Cod in the inshore and offshore waters of Labrador and northeastern Newfoundland, including Grand Bank, having declined 97% since the early 1970s and more than 99% since the early 1960s, are now at historically low levels. -

Hypotheses for the Decline of Cod in the North Atlantic*
MARINE ECOLOGY PROGRESS SERIES Vol. 138: 293-308, 1996 Published July 25 Mar Ecol Prog Ser REVIEW Hypotheses for the decline of cod in the North Atlantic* Ransom A. ~yers'#**,Jeffrey A. ~utchings~,N. J. Barrowman' 'Northwest Atlantic Fisheries Centre, Science Branch, PO Box 5667, St. John's, Newfoundland, Canada AlC 5x1 'Department of Biology, Dalhousie University, Halifax, Nova Scotia, Canada B3H 451 ABSTRACT We examine alternative hypotheses for the decllne of 20 cod Gadus morhua stocks in the North Atlantlc The year of the lowest observed biomass of spawners did not correspond to low juve- nile survival for the cohorts that should have contnbuted to the stock in that year However, fishing mortdl~tywas very high for the years preceding the collapse The collapse of the cod stocks was not caused by a lack of resilience at low population abundance because all spawners were able to produce many potential replacements at low population size We show that as populations collapsed, flshlng mortality increased untll the populations were reduced to very low levels We conclude that increased fishing mortality caused the population decl~nes,and often the collapses, of the cod stocks KEY WORDS Gadus morhua Cod North Atlantic Decline Stocks Tlme serles Spawners Recru~tment Catch Mortality Collapse Density-dependent mortality Fishlng INTRODUCTION a fishery. We have reformulated these questions as follows: During the last few years many of the world's cod (1)What was the extent and timing of the population Gadus morhua stocks have rapidly declined to the decline? point where fishing has been effectively eliminated. -

The Commonwealth of Massachusetts Division of Marine Fisheries
The Commonwealth of Massachusetts Division of Marine Fisheries 251 Causeway Street, Suite 400, Boston, MA 02114 p: (617) 626-1520 | f: (617) 626-1509 www.mass.gov/marinefisheries CHARLES D. BAKER KARYN E. POLITO KATHLEEN A. THEOHARIDES RONALD S. AMIDON DANIEL J. MCKIERNAN Governor Lt. Governor Secretary Commissioner Director February 19, 2021 MarineFisheries Advisory New Protected Species Regulations Finalized for Fixed Gear Fisheries and Industry Outreach on Required Gear Modifications This advisory serves to provide you with information regarding new protected species regulations. This includes a description of the new regulatory requirements, resources available to assist commercial trap fishermen in complying with the new gear modification and weak rope rules, and additional information germane to DMF’s ongoing efforts to protect the North Atlantic right whale. New Regulations Enacted The Marine Fisheries Advisory Commission approved several new regulatory measures at their January 28, 2021 business meeting affecting protected species and fixed gear fishing in Massachusetts (Advisory). These regulations have been filed with the Secretary of State and will be implemented as of March 5, 2021. The seasonal closure rules will be effective March 5, 2021, whereas the gear modification rules will not go into effect until May 2021 to provide fishermen with time to alter their gear configuration. More details on each new regulatory provision are provided in the bullets below. • Seasonal Commercial Trap Gear Closure. A new commercial trap gear closure goes into effect on March 5, 2021. This new regulation extends the prior seasonal state waters commercial trap gear closure in both space and time. The prior closure occurred from February 1 – April 30 within Cape Cod Bay, Stellwagen Bank, and the Outer Cape Cod Lobster Management Area. -

Failing Fish
Failing Fish ----Advertisement---- ----Advertisement---- HOME Failing Fish NEWS COMMENTARY News: A sampling of creatures at serious risk of disappearing from our oceans and our dinner plates ARTS MOJOBLOG Illustrations by Jack Unruh RADIO CUSTOMER March/April 2006 Issue SERVICE DONATE STORE ABOUT US NEWSLETTERS SUBSCRIBE ADVERTISE Bluefin Tuna Warm-blooded bluefins, which can weigh 1,500 punds, are one of the largest bony fish swimming the seas. The Atlantic bluefin population has fallen by more than 80 percent since the 1970s; Pacific stocks are also dwindling. Advanced Search Browse Back Issues http://www.motherjones.com/news/feature/2006/03/failing_fish.html (1 of 4)2/23/2006 1:30:09 PM Failing Fish Read the Current Issue BUY THIS ISSUE SUBSCRIBE NOW Blue Crab Since Chesapeake Bay harvests are half of what they were a decade ago, at least 70 percent of crabmeat CRAZY PRICE! products sold in the United States now contain foreign crabs. 1 year just $10 Click Here Sundays on Air America Radio THIS WEEK The roots of the Eastern Oyster conflict over the Ships in the Chesapeake Bay once had to steer around massive oyster reefs. Poor water quality, exotic Danish Mohammed parasites, and habitat destruction have reduced the Chesapeake oyster stock to 1 percent of its historic level. cartoons, Clinton's economic advisor on Bush's troubles, and Iraq war veterans running for office as Democrats..... Learn More... Blue Marlin Since longlines replaced harpoons in the early 1960s, the Atlantic blue marlin has been driven toward extinction. A quarter of all blue marlin snared by longlines are dead by the time they reach the boat. -

The Political Ecology of Crisis and Institutional Change: the Case of the Northern Cod1
The Political Ecology of Crisis and Institutional Change: 1 The Case of the Northern Cod by Bonnie J. McCay and Alan Christopher Finlayson Rutgers the State University, New Brunswick, New Jersey Presented to the Annual Meetings of the American Anthropological Association, Washington, D.C., November 15-19, 1995. A longer and different version, with Finlayson as first author, is to be published in Linking Social and Ecological Systems; Institutional Learning for Resilience. Fikret Berkes and Carl Folke, eds. on behalf of The Property Rights Programme, The Beijer International Institute of Ecological Economics, The Royal Swedish Academy of Sciences. Introduction: Systems Crisis The question we pose, although cannot yet answer with certainty, is whether the current set of crises in fisheries--- from the salmon of the West Coast of North America to the groundfish of the East Coast of North America--will open the door to institutional- cultural, social, political- change. There are theoretical and logical grounds for thinking that crisis and institutional change might be linked (Holling 1986; Kuhn 1962, 1970) although the nature and outcomes of linkages are not so straightforward (Lee 1993). Fisheries management is a thoroughly modernist venture, imbued as are so many other of the applied "natural resource" areas with a very pragmatic, utilitarian, science- dependent, and mostly optimistic perspective on the ability of people to "manage" wild things and processes. Management decisions-such as quotas, the timing or length of a fishing season, and the kinds of fishing gear allowed-are influenced by many things but are in theory dependent on information and understandings from a probabilistic but deterministic science known as "stock assessment." However, the alarming occurrence of fish stock declines and collapses throughout the world is accompanied by critical appraisals of the specific tools and the more general philosophies of fisheries management. -

Pollachius Virens
MARINE ECOLOGY PROGRESS SERIES Published October 5 Mar Ecol Prog Ser Use of rocky intertidal habitats by juvenile pollock Pollachius virens Robert W. Rangeley*, Donald L. Kramer Department of Biology, McGill University, 1205 Docteur Penfield Avenue, Montreal, Quebec, Canada H3A 1B1 ABSTRACT: We ~nvestigatedpatterns of distribution and foraging by young-of-the-year pollock Pol- lachius virens in the rocky intertidal zone. Pollock were sampled by beach seine in fucoid macroalgae and in open habitats at all stages of the tide, day and night throughout the summer. Their presence in shallow water at the high tidal stages indicated that at least part of the pollock population migrated across the full width of the intertidal zone (150 m) each tide. Densities in shallow water were much higher at low than at high tidal stages suggesting that a large influx of pollock moved in from the sub- tidal zone at low tidal stages and then dispersed into intertidal habitats at high tidal stages. There were few differences in pollock densit~esbetween algal and open habitats but abundances likely increased in the algal habitat at higher tidal stages when changes in habitat availability are taken Into account. Densities were higher at night and there was an order of magnitude decline in pollock densities from early to late summer. In another study we showed that piscivorous birds are a probable cause of pollock summer mortality. Pollock fed on invertebrates from intertidal algae relatively continuously. The tidal migrations of juvenile pollock observed in this study and their use of macroalgae as a foraging and possibly a refuging habitat strongly suggests that the rocky intertidal zone may be an important fish nursery area.