The Knight and the King
Total Page:16
File Type:pdf, Size:1020Kb

Load more
Recommended publications
-

Civil Aviation Development Investment Program (Tranche 3)
Resettlement Due Diligence Reports Project Number: 43141-044 June 2016 PNG: Multitranche Financing Facility - Civil Aviation Development Investment Program (Tranche 3) Prepared by National Airports Corporation for the Asian Development Bank. This resettlement due diligence report is a document of the borrower. The views expressed herein do not necessarily represent those of ADB's Board of Directors, Management, or staff, and may be preliminary in nature. Your attention is directed to the “terms of use” section of this website. In preparing any country program or strategy, financing any project, or by making any designation of or reference to a particular territory or geographic area in this document, the Asian Development Bank does not intend to make any judgments as to the legal or other status of any territory or area. Table of Contents B. Resettlement Due Diligence Report 1. Madang Airport Due Diligence Report 2. Mendi Airport Due Diligence Report 3. Momote Airport Due Diligence Report 4. Mt. Hagen Due Diligence Report 5. Vanimo Airport Due Diligence Report 6. Wewak Airport Due Diligence Report 4. Madang Airport Due Diligence Report. I. OUTLINE FOR MADANG AIRPORT DUE DILIGENCE REPORT 1. The is a Due Diligent Report (DDR) that reviews the Pavement Strengthening Upgrading, & Associated Works proposed for the Madang Airport in Madang Province (MP). It presents social safeguard aspects/social impacts assessment of the proposed works and mitigation measures. II. BACKGROUND INFORMATION 2. Madang Airport is situated at 5° 12 30 S, 145° 47 0 E in Madang and is about 5km from Madang Town, Provincial Headquarters of Madang Province where banks, post office, business houses, hotels and guest houses are located. -

Health&Medicalinfoupdate8/10/2017 Page 1 HEALTH and MEDICAL
HEALTH AND MEDICAL INFORMATION The American Embassy assumes no responsibility for the professional ability or integrity of the persons, centers, or hospitals appearing on this list. The names of doctors are listed in alphabetical, specialty and regional order. The order in which this information appears has no other significance. Routine care is generally available from general practitioners or family practice professionals. Care from specialists is by referral only, which means you first visit the general practitioner before seeing the specialist. Most specialists have private offices (called “surgeries” or “clinic”), as well as consulting and treatment rooms located in Medical Centers attached to the main teaching hospitals. Residential areas are served by a large number of general practitioners who can take care of most general illnesses The U.S Government assumes no responsibility for payment of medical expenses for private individuals. The Social Security Medicare Program does not provide coverage for hospital or medical outside the U.S.A. For further information please see our information sheet entitled “Medical Information for American Traveling Abroad.” IMPORTANT EMERGENCY NUMBERS AMBULANCE/EMERGENCY SERVICES (National Capital District only) Police: 112 / (675) 324-4200 Fire: 110 St John Ambulance: 111 Life-line: 326-0011 / 326-1680 Mental Health Services: 301-3694 HIV/AIDS info: 323-6161 MEDEVAC Niugini Air Rescue Tel (675) 323-2033 Fax (675) 323-5244 Airport (675) 323-4700; A/H Mobile (675) 683-0305 Toll free: 0561293722468 - 24hrs Medevac Pacific Services: Tel (675) 323-5626; 325-6633 Mobile (675) 683-8767 PNG Wide Toll free: 1801 911 / 76835227 – 24hrs Health&MedicalInfoupdate8/10/2017 Page 1 AMR Air Ambulance 8001 South InterPort Blvd Ste. -

First Records of Dragonflies (Odonata) from the Foja Mountains, Papua Province, Indonesia
14 Suara Serangga Papua, 2009, 4 (1) Juli- September 2009 First records of dragonflies (Odonata) from the Foja Mountains, Papua Province, Indonesia 1 2 Vincent J. Kalkman , Henk van Mastrigt & Stephen J. Rlchards" 1Nationaal Natuurhistorisch Museum - Naturalis Postbus 9517, NL-2300 RA Leiden, THE NETHERLANDS Email: [email protected] 2 Kelompok Entomologi Papua, Kotakpos 1078, Jayapura 99010, Papua, INDONESIA Email: [email protected] 3 Vertebrates Department, South Australian Museum, North Terrace, Adelaide, SA 5000, AUSTRALlA and Rapid Assessment Program, Conservation International, Atherton, Queensland 4883, AUSTRALlA Email: [email protected] Suara Serangga Papua: 4 (1): 14 - 19 Abstract: A small collection of dragonflies obtained during two RAP biodiversity surveys to the Foja Mountains, organised by Conservation International with help of LlPI, Bogor, in 2005 and 2008 are brought on record. Twelve species were found at two sites below 100 m near Kwerba, a small village adjacent to the Mamberamo River. Thirteen species were recorded at 'Moss Camp' at 1650 m in the Foja Mountains. Of these Hemicordulia ericetorum was previously only known from the central mountain range while Oreaeschna dictatrix was only known from Lake Paniai and the Cyclops Mountains. It is likely that more genera and species now known onlyfrom the central mountain range occur in the Foja Mountains and probably also the Van Rees Mountains. However one species, Argiolestes spec. nov. is probably endemie to the Foja Mountains. Although this collection includes only a small fraction of the diversity likely to be present in the mountains it is nonetheless of interest as it represents the first records of dragonflies from the area. -

Coronavirus Covid-19 in Papua New Guinea
Coronavirus Covid-19 in Papua New Guinea Bryant Allen In Brief 2020/6 Introduction distances on foot but the infectious agents were moved from The first coronavirus Covid-19 case entered Papua New Guinea person to person and from village to village, over long distances. on 13 March 2020. Whether it will spread is a critical question, The Highlands dysentery spread in spite of the army and patrol but it is likely. This In Brief examines two previous epidemics in officers attempting to stop people moving. PNG and then describes the population and the movements of Covid-19 in PNG people who will carry the virus with them, to suggest how this epidemic will develop. It was inevitable that the Covid-19 virus would enter PNG with an infected person arriving from overseas. It was predictable they Smallpox and dysentery would arrive at Port Moresby, a city of at least 350,000 people. In 1890 a viral smallpox was introduced into a German colonial On 22 March 2020 seven international flights arrived there with outpost just south of Madang from a German ship (Sack and more than 1600 passengers, 62 per cent from Brisbane and Clarke 1979:81). Smallpox spread as far west as Nuku District Cairns and 30 per cent from Singapore and Manila. But the in Sandaun Province and across Vitiaz Strait as far east as origins of the flights do not provide information about where Talasea District in West New Britain. It may have spread further the passengers had been in the previous 14 days. The infected but oral historical accounts have not been collected. -

Papua New Guinea (And Comparators)
Public Disclosure Authorized Public Disclosure Authorized Public Disclosure Authorized Public Disclosure Authorized BELOW THE GLASS FLOOR Analytical Review of Expenditure by Provincial Administrations on Rural Health from Health Function Grants & Provincial Internal Revenue JULY 2013 Below the Glass Floor: An Analytical Review of Provincial Administrations’ Rural Health Expenditure Rights and Permissions The material in this publication is copyrighted. Copying and/or transmitting portions or all of this work without permission may be a violation of applicable law. The International Bank for Reconstruction and Development/ The World Bank encourages dissemination of its work and will normally grant permission to reproduce portions of the work promptly. For permission to photocopy or reprint any part of this work, please send a request with complete information to the Copyright Clearance Center Inc., 222 Rosewood Drive, Danvers, MA 01923, USA; telephone: 978-750- 8400; fax: 978-750-4470; Internet: www.copyright.com. All other queries on rights and licenses, including subsidiary rights, should be addressed to the Office of the Publisher, The World Bank, 1818 H Street, NW, Washington, DC 20433, USA; fax 202-522-2422; email: [email protected]. - i - Below the Glass Floor: An Analytical Review of Provincial Administrations’ Rural Health Expenditure Table of Contents Acknowledgment ......................................................................................................................... 5 Abbreviations ............................................................................................................................. -

Two New Frog Species from the Foja Mountains in Northwestern New Guinea (Amphibia, Anura, Microhylidae)
68 (2): 109 –122 © Senckenberg Gesellschaft für Naturforschung, 2018. 28.5.2018 Two new frog species from the Foja Mountains in north western New Guinea (Amphibia, Anura, Micro hylidae) Rainer Günther 1, Stephen Richards 2 & Burhan Tjaturadi 3 1 Museum für Naturkunde, Invalidenstr. 43, 10115 Berlin, Germany; [email protected] — 2 Herpetology Department, South Australian Museum, North Terrace, Adelaide, South Australia 5000, Australia; [email protected] — 3 Conservation Inter- national – Papua Program. Current address: Center for Environmental Studies, Sanata Dharma University (CESSDU), Yogyakarta, Indonesia; [email protected] Accepted January 18, 2018. Published online at www.senckenberg.de/vertebrate-zoology on May 28, 2018. Editor in charge: Raffael Ernst Abstract Two new microhylid frogs in the genera Choerophryne and Oreophryne are described from the Foja Mountains in Papua Province of Indonesia. Both are small species (males 15.9 – 18.5 mm snout-urostyle length [SUL] and 21.3 – 22.9 mm SUL respectively) that call from elevated positions on foliage in primary lower montane rainforest. The new Choerophryne species can be distinguished from all congeners by, among other characters, a unique advertisement call consisting of an unpulsed (or very finely pulsed) peeping note last- ing 0.29 – 0.37 seconds. The new Oreophryne species belongs to a group that has a cartilaginous connection between the procoracoid and scapula and rattling advertisement calls. Its advertisement call is a loud rattle lasting 1.2 – 1.5 s with a note repetition rate of 11.3 – 11.7 notes per second. Kurzfassung Es werden zwei neue Engmaulfrösche der Gattungen Choerophryne und Oreophryne aus den Foja-Bergen in der Papua Provinz von Indonesien beschrieben. -

RAPID ASSESSMENT of AVOIDABLE BLINDNESS and DIABETIC RETINOPATHY REPORT Papua New Guinea 2017
RAPID ASSESSMENT OF AVOIDABLE BLINDNESS AND DIABETIC RETINOPATHY REPORT Papua New Guinea 2017 RAPID ASSESSMENT OF AVOIDABLE BLINDNESS AND DIABETIC RETINOPATHY PAPUA NEW GUINEA, 2017 1 Acknowledgements The Rapid Assessment of Avoidable Blindness (RAAB) + Diabetic Retinopathy (DR) was a Brien Holden Vision Institute (the Institute) project, conducted in cooperation with the Institute’s partner in Papua New Guinea (PNG) – PNG Eye Care. We would like to sincerely thank the Fred Hollows Foundation, Australia for providing project funding, PNG Eye Care for managing the field work logistics, Fred Hollows New Zealand for providing expertise to the steering committee, Dr Hans Limburg and Dr Ana Cama for providing the RAAB training. We also wish to acknowledge the National Prevention of Blindness Committee in PNG and the following individuals for their tremendous contributions: Dr Jambi Garap – President of National Prevention of Blindness Committee PNG, Board President of PNG Eye Care Dr Simon Melengas – Chief Ophthalmologist PNG Dr Geoffrey Wabulembo - Paediatric ophthalmologist, University of PNG and CBM Mr Samuel Koim – General Manager, PNG Eye Care Dr Georgia Guldan – Professor of Public Health, Acting Head of Division of Public Health, School of Medical and Health Services, University of PNG Dr Apisai Kerek – Ophthalmologist, Port Moresby General Hospital Dr Robert Ko – Ophthalmologist, Port Moresby General Hospital Dr David Pahau – Ophthalmologist, Boram General Hospital Dr Waimbe Wahamu – Ophthalmologist, Mt Hagen Hospital Ms Theresa Gende -

Building Capacity for Multidisciplinary Landscape Assessment in Papua: Three Phases of Training and Pilot Assessments in the Mamberamo Basin
P.O. Box 6596 JKPWB Tel. (62) 251 622622 Jakarta 10065 Fax (62) 251 622100 CIFOR Indonesia Building Capacity for Multidisciplinary Landscape Assessment in Papua: three phases of training and pilot assessments in the Mamberamo Basin A report based on work jointly undertaken in 2004 by the Center for International Forestry Research, Conservation International (Papua Program), Lembaga Ilmu Pengetahuan Indonesia (LIPI), 18 Papua- based Trainees, and the people of Papasena-I and Kwerba Villages. MLA in Papua, Page 1 SUMMARY Conservation International (CI) supports a number of ongoing initiatives in the Mamberamo area of Papua. The principal aims are to strengthen biodiversity conservation and environmental management and facilitate the creation of a ‘Mamberamo Biodiversity Conservation Corridor’, which links currently established protected areas through strategically placed ‘indigenous forest reserves’. Two primary requirements are 1) to find suitable means to allow local communities to participate in decision-making processes, and 2) capacity building of locally based researchers to assist in planning and developing this program. The MLA training reported here is designed to build capacity and assess options and opportunities within this context. The Centre for International Forestry Research (CIFOR) has developed methods for assessing 'what really matters' to communities living in tropical forest landscapes. Known as the Multidisciplinary Landscape Assessment or ‘MLA' (see http://www.cifor.cgiar.org/mla), this approach enhances understanding amongst conservation and development practitioners, policy makers and forest communities. Information yielded through the MLA can identify where local communities’ interests and priorities might converge (or conflict) with conservation and sustainable development goals. CIFOR's MLA methods have already been applied in Indonesia (East Kalimantan), Bolivia and Cameroon. -
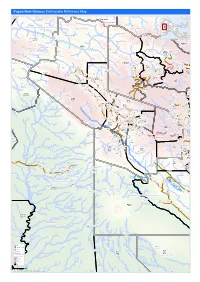
Earthquake Reference Map
Papua New Guinea: Earthquake Reference Map Telefomin London Rural Lybia Telefomin Tunap/Hustein District Ambunti/Drekikier District Kaskare Akiapjmin WEST SEPIK (SANDAUN) PROVINCE Karawari Angoram EAST SEPIK PROVINCE Rural District Monduban HELA PROVINCE Wundu Yatoam Airstrip Lembana Kulupu Malaumanda Kasakali Paflin Koroba/Kopiago Kotkot Sisamin Pai District Emo Airstrip Biak Pokale Oksapmin Liawep Wabag Malandu Rural Wayalima District Hewa Airstrip Maramuni Kuiva Pauteke Mitiganap Teranap Lake Tokom Rural Betianap Puali Kopiago Semeti Oksapmin Sub Aipaka Yoliape Waulup Oksapmin District ekap Airport Rural Kenalipa Seremty Divanap Ranimap Kusanap Tomianap Papake Lagaip/Pogera Tekin Winjaka Airport Gawa Eyaka Airstrip Wane 2 District Waili/Waki Kweptanap Gaua Maip Wobagen Wane 1 Airstrip aburap Muritaka Yalum Bak Rural imin Airstrip Duban Yumonda Yokona Tili Kuli Balia Kariapuka Yakatone Yeim Umanap Wiski Aid Post Poreak Sungtem Walya Agali Ipate Airstrip Piawe Wangialo Bealo Paiela/Hewa Tombaip Kulipanda Waimalama Pimaka Ipalopa Tokos Ipalopa Lambusilama Rural Primary Tombena Waiyonga Waimalama Taipoko School Tumundane Komanga ANDS HIG PHaLin Yambali Kolombi Porgera Yambuli Kakuane C/Mission Paiela Aspiringa Maip Pokolip Torenam ALUNI Muritaka Airport Tagoba Primary Kopetes Yagoane Aiyukuni SDA Mission Kopiago Paitenges Haku KOPIAGO STATION Rural Lesai Pali Airport School Yakimak Apostolic Mission Politika Tamakale Koemale Kambe Piri Tarane Pirika Takuup Dilini Ingilep Kiya Tipinini Koemale Ayene Sindawna Taronga Kasap Luth. Yaparep -

From the Foja Mountains, Papua, Indonesia (Lepidoptera: Pieridae)
Suara Serangga Papua, 2009, 3 (3) Januari - Maret 2009 SOME Notes ON Delias (Hübner, 1819) from the Foja Mountains, Papua, Indonesia (Lepidoptera: PIeridae) HENK vaN Mastrigt Kelompok Entomologi Papua, Kotakpos 1078, Jayapura 99010, INDONESIA E.mail: [email protected];[email protected] Suara Serangga Papua 3(3): 1 - 13 Abstract: The second survey to the Foja Mts (at 1,650 m) increased the number of Delias species recorded in th at area from eight to twelve, including a new species described below. On 1,250 m three species were collected, including one not recorded at 1,650 m. Further information about the Foja Delias, including descriptions of the female of D. durai and the male of D. microsticha weja is provided. /khtisar: Survei kedua ke Pegunungan Foja meningkatkan jumlah spesies De/ias di daerah itu (1.650 m) dari delapan menjadi dua belas, termasuk satu spesies baru yang diletakkan di bawah ini. Pada ketinggian 1.250 m tiga spesies ditangkap, termasuk satu yang tidak diobservasi pada ketinggian 1.650 m. Catatan-catatan tambahan diberikan tentang Delias dari Peg. Foja, termasuk deskripsi betina D. durai dan jantan D. microsticha weja. Keywords: new species. Introduction After the first successful survey of the Foja Mountains in the northern part of Papua, close to the Mamberamo River, (25 November - 7 December 2005) many of the participants hoped to return to the isolated mountains range once more. In November 2008 this became possible. Conservation International in collaboration with the Zoological Division of the Research Center for Biology of the Indonesian Institute of Science (LIP!)organized a survey to Kwerba and from there by helicopter to the Foja Mountains. -

Three New Species of Argio/Estes, with a Key to the Males of Argiolestes S
Received 16 July 2009; revised and accepted 13 February 2010 Three new species of Argio/estes, with a key to the males of Argiolestes s. str. (Odonata: Megapodagrionidae) 1 2 3 Vincent J. Kalkman , Stephen J. Richards & Dan A. Polhemus 1 National Museum of Natural History, P.O. Box 9517, 2300 RA Lei den, The Netherlands. <[email protected]> 2Vertebrates Department, South Australian Museum, North Terrace, Adelaide, SA 5000, Australia. <[email protected]> 3 Department of Natural Sciences, Bishop Museum, 1525 Ben ice St., Honolulu, Hl96817, USA. <[email protected]> Keywords: Odonata, dragonfly, Zygoptera, Megapodagrionidae, Argialestes, taxonomy, New Guinea, Sulawesi, Moluccas. ABSTRACT In this article the genus Argialestes s.str. is defined and three new species belonging to this group are described: A. faja sp. nov. (holotype: Foja Mountains, Indonesia, dep. in MBBJ); A. muller sp. nov. (holotype: Baia River, Papua New Guinea, dep. in SAMA); A. roan sp. nov. (holotype: Roon Island, Indonesia, dep. in BPBM). New records for A. alfurus are given, a key to males is presented and a map of the distribution of the species is shown. Both sexes of A. muller and the male of A. roan are depicted in life. The group is distributed from New Guinea over the Moluccas to Sulawesi. As far as is known all species are confined to forest brooks. INTRODUCTION The genus Argialestes currently includes 48 species found on New Caledonia, the Solomon Islands, the Bismarck Archipelago, New Guinea and adjacent smaller islands, the Moluccas, Sulawesi and Luzon. Most of these were described by Lief tinck (1935, 1938, 1949, 1956) but recently, renewed interest in the genus has led to the description of several additional species (Englund & Polhemus 2007; Kalk man 2007, 2008a; Theischinger & Richards 2006, 2007, 2008; Michalski & Oppel 2010). -

Estimating Adult Mortality in Papua New Guinea, 2011 Urarang Kitur* , Tim Adair and Alan D
Kitur et al. Population Health Metrics (2019) 17:4 https://doi.org/10.1186/s12963-019-0184-x RESEARCH Open Access Estimating Adult Mortality in Papua New Guinea, 2011 Urarang Kitur* , Tim Adair and Alan D. Lopez Abstract Background: Mortality in Papua New Guinea (PNG) is poorly measured because routine reporting of deaths is incomplete and inaccurate. This study provides the first estimates in the academic literature of adult mortality (45q15) in PNG by province and sex. These results are compared to a Composite Index of provincial socio- economic factors and health access. Methods: Adult mortality estimates (45q15) by province and sex were derived using the orphanhood method from data reported in the 2000 and 2011 national censuses. Male adult mortality was adjusted based on the estimated incompleteness of mortality reporting. The Composite Index was developed using the mean of education, economic and health access indicators from various data sources. Results: Adult mortality for PNG in 2011 was estimated as 269 per 1000 for males and 237 for females. It ranged from 197 in Simbu to 356 in Sandaun province among men, and from 164 in Western Highlands to 326 in Gulf province among women. Provinces with a low Composite Index (Sandaun, Gulf, Enga and Southern Highlands) had comparatively high levels of adult mortality for both sexes, while provinces with a higher Composite Index (National Capital District and Manus) reported lower adult mortality. Conclusions: Adult mortality in PNG remains high compared with other developing countries. Provincial variations in mortality correlate with the Composite Index. Health and development policy in PNG needs to urgently address the main causes of persistent high premature adult mortality, particularly in less developed provinces.