Human Visual Perception
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

MITOCW | 9: Receptive Fields - Intro to Neural Computation
MITOCW | 9: Receptive Fields - Intro to Neural Computation MICHALE FEE: So today, we're going to introduce a new topic, which is related to the idea of fine- tuning curves, and that is the notion of receptive fields. So most of you have probably been, at least those of you who've taken 9.01 or 9.00 maybe, have been exposed to the idea of what a receptive field is. The idea is basically that in sensory systems neurons receive input from the sensory periphery, and neurons generally have some kind of sensory stimulus that causes them to spike. And so one of the classic examples of how to find receptive fields comes from the work of Huble and Wiesel. So I'll show you some movies made from early experiments of Huble-Wiesel where they are recording in the visual cortex of the cat. So they place a fine metal electrode into a primary visual cortex, and they present. So then they anesthetize the cat so the cat can't move. They open the eye, and the cat's now looking at a screen that looks like this, where they play a visual stimulus. And they actually did this with essentially a slide projector that they could put a card in front of that had a little hole in it, for example, that allowed a spot of light to project onto the screen. And then they can move that spot of light around while they record from neurons in visual cortex and present different visual stimuli to the retina. -

The Roles and Functions of Cutaneous Mechanoreceptors Kenneth O Johnson
455 The roles and functions of cutaneous mechanoreceptors Kenneth O Johnson Combined psychophysical and neurophysiological research has nerve ending that is sensitive to deformation in the resulted in a relatively complete picture of the neural mechanisms nanometer range. The layers function as a series of of tactile perception. The results support the idea that each of the mechanical filters to protect the extremely sensitive recep- four mechanoreceptive afferent systems innervating the hand tor from the very large, low-frequency stresses and strains serves a distinctly different perceptual function, and that tactile of ordinary manual labor. The Ruffini corpuscle, which is perception can be understood as the sum of these functions. located in the connective tissue of the dermis, is a rela- Furthermore, the receptors in each of those systems seem to be tively large spindle shaped structure tied into the local specialized for their assigned perceptual function. collagen matrix. It is, in this way, similar to the Golgi ten- don organ in muscle. Its association with connective tissue Addresses makes it selectively sensitive to skin stretch. Each of these Zanvyl Krieger Mind/Brain Institute, 338 Krieger Hall, receptor types and its role in perception is discussed below. The Johns Hopkins University, 3400 North Charles Street, Baltimore, MD 21218-2689, USA; e-mail: [email protected] During three decades of neurophysiological and combined Current Opinion in Neurobiology 2001, 11:455–461 psychophysical and neurophysiological studies, evidence has accumulated that links each of these afferent types to 0959-4388/01/$ — see front matter © 2001 Elsevier Science Ltd. All rights reserved. a distinctly different perceptual function and, furthermore, that shows that the receptors innervated by these afferents Abbreviations are specialized for their assigned functions. -

Auditory Experience Controls the Maturation of Song Discrimination and Sexual Response in Drosophila Xiaodong Li, Hiroshi Ishimoto, Azusa Kamikouchi*
RESEARCH ARTICLE Auditory experience controls the maturation of song discrimination and sexual response in Drosophila Xiaodong Li, Hiroshi Ishimoto, Azusa Kamikouchi* Graduate School of Science, Nagoya University, Nagoya, Japan Abstract In birds and higher mammals, auditory experience during development is critical to discriminate sound patterns in adulthood. However, the neural and molecular nature of this acquired ability remains elusive. In fruit flies, acoustic perception has been thought to be innate. Here we report, surprisingly, that auditory experience of a species-specific courtship song in developing Drosophila shapes adult song perception and resultant sexual behavior. Preferences in the song-response behaviors of both males and females were tuned by social acoustic exposure during development. We examined the molecular and cellular determinants of this social acoustic learning and found that GABA signaling acting on the GABAA receptor Rdl in the pC1 neurons, the integration node for courtship stimuli, regulated auditory tuning and sexual behavior. These findings demonstrate that maturation of auditory perception in flies is unexpectedly plastic and is acquired socially, providing a model to investigate how song learning regulates mating preference in insects. DOI: https://doi.org/10.7554/eLife.34348.001 Introduction Vocal learning in infants or juvenile birds relies heavily on the early experience of the adult conspe- cific sounds. In humans, early language input is necessary to form the ability of phonetic distinction *For correspondence: and pattern detection in the phase of auditory learning (Doupe and Kuhl, 1999; Kuhl, 2004). [email protected] Because of the strong parallels between speech acquisition of humans and song learning of song- Competing interests: The birds, and the difficulties to investigate the neural mechanisms of human early auditory memory at authors declare that no cellular resolution, songbirds have been used as a predominant model in studying memory formation competing interests exist. -

Center Surround Receptive Field Structure of Cone Bipolar Cells in Primate Retina
Vision Research 40 (2000) 1801–1811 www.elsevier.com/locate/visres Center surround receptive field structure of cone bipolar cells in primate retina Dennis Dacey a,*, Orin S. Packer a, Lisa Diller a, David Brainard b, Beth Peterson a, Barry Lee c a Department of Biological Structure, Uni6ersity of Washington, Box 357420, Seattle, WA 98195-7420, USA b Department of Psychology, Uni6ersity of California Santa Barbara, Santa Barbara, CA, USA c Max Planck Institute for Biophysical Chemistry, Gottingen, Germany Received 28 September 1999; received in revised form 5 January 2000 Abstract In non-mammalian vertebrates, retinal bipolar cells show center-surround receptive field organization. In mammals, recordings from bipolar cells are rare and have not revealed a clear surround. Here we report center-surround receptive fields of identified cone bipolar cells in the macaque monkey retina. In the peripheral retina, cone bipolar cell nuclei were labeled in vitro with diamidino-phenylindole (DAPI), targeted for recording under microscopic control, and anatomically identified by intracellular staining. Identified cells included ‘diffuse’ bipolar cells, which contact multiple cones, and ‘midget’ bipolar cells, which contact a single cone. Responses to flickering spots and annuli revealed a clear surround: both hyperpolarizing (OFF) and depolarizing (ON) cells responded with reversed polarity to annular stimuli. Center and surround dimensions were calculated for 12 bipolar cells from the spatial frequency response to drifting, sinusoidal luminance modulated gratings. The frequency response was bandpass and well fit by a difference of Gaussians receptive field model. Center diameters were all two to three times larger than known dendritic tree diameters for both diffuse and midget bipolar cells in the retinal periphery. -
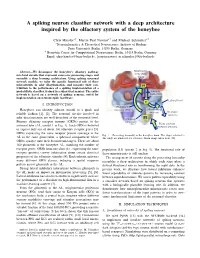
A Spiking Neuron Classifier Network with a Deep Architecture Inspired By
A spiking neuron classifier network with a deep architecture inspired by the olfactory system of the honeybee Chris Hausler¨ ∗†, Martin Paul Nawrot∗† and Michael Schmuker∗† ∗Neuroinformatics & Theoretical Neuroscience, Institute of Biology Freie Universitat¨ Berlin, 14195 Berlin, Germany † Bernstein Center for Computational Neuroscience Berlin, 10115 Berlin, Germany Email: [email protected], fmartin.nawrot, [email protected] Abstract—We decompose the honeybee’s olfactory pathway Mushroom into local circuits that represent successive processing stages and body (MB) calyx resemble a deep learning architecture. Using spiking neuronal network models, we infer the specific functional role of these microcircuits in odor discrimination, and measure their con- 2 tribution to the performance of a spiking implementation of a probabilistic classifier, trained in a supervised manner. The entire Kenyon network is based on a network of spiking neurons, suited for Cells (KC) implementation on neuromorphic hardware. Lateral horn 3 I. INTRODUCTION Projection Honeybees can identify odorant stimuli in a quick and neurons (PN) MB-extrinsic neurons To motor reliable fashion [1], [2]. The neuronal circuits involved in neurons odor discrimination are well described at the structural level. 1 Antennal Primary olfactory receptor neurons (ORNs) project to the lobe (AL) From odorant antennal lobe (AL, circuit 1 in Fig. 1). Each ORN is believed receptor neurons to express only one of about 160 olfactory receptor genes [3]. ORNs expressing the same receptor protein converge in the Fig. 1. Processing hierarchy in the honeybee brain. The stages relevant to AL in the same glomerulus, a spherical compartment where this study are numbered for reference. -

The Visual System: Higher Visual Processing
The Visual System: Higher Visual Processing Primary visual cortex The primary visual cortex is located in the occipital cortex. It receives visual information exclusively from the contralateral hemifield, which is topographically represented and wherein the fovea is granted an extended representation. Like most cortical areas, primary visual cortex consists of six layers. It also contains, however, a prominent stripe of white matter in its layer 4 - the stripe of Gennari - consisting of the myelinated axons of the lateral geniculate nucleus neurons. For this reason, the primary visual cortex is also referred to as the striate cortex. The LGN projections to the primary visual cortex are segregated. The axons of the cells from the magnocellular layers terminate principally within sublamina 4Ca, and those from the parvocellular layers terminate principally within sublamina 4Cb. Ocular dominance columns The inputs from the two eyes also are segregated within layer 4 of primary visual cortex and form alternating ocular dominance columns. Alternating ocular dominance columns can be visualized with autoradiography after injecting radiolabeled amino acids into one eye that are transported transynaptically from the retina. Although the neurons in layer 4 are monocular, neurons in the other layers of the same column combine signals from the two eyes, but their activation has the same ocular preference. Bringing together the inputs from the two eyes at the level of the striate cortex provide a basis for stereopsis, the sensation of depth perception provided by binocular disparity, i.e., when an image falls on non-corresponding parts of the two retinas. Some neurons respond to disparities beyond the plane of fixation (far cells), while others respond to disparities in front of the plane of the fixation (near cells). -

The Retina and Vision
CHAPTER 19 The Retina and Vision The visual system is arguably the most important system through which our brain gathers information about our surroundings, and forms one of our most complex phys- iological systems. In vertebrates, light entering the eye through the lens is detected by photosensitive pigment in the photoreceptors, converted to an electrical signal, and passed back through the layers of the retina to the optic nerve, and from there, through the visual nuclei, to the visual cortex of the brain. At each stage, the signal passes through an elaborate system of biochemical and neural feedbacks, the vast majority of which are poorly, if at all, understood. Although there is great variety in detail between the eyes of different species, a number of important features are qualitatively conserved. Perhaps the most striking of these features is the ability of the visual system to adapt to background light. As the background light level increases, the sensitivity of the visual system is decreased, which allows for operation over a huge range of light levels. From a dim starlit night to a bright sunny day, the background light level varies over 10 orders of magnitude (Hood and Finkelstein, 1986), and yet our eyes continue to operate across all these levels without becoming saturated with light. The visual system accomplishes this by ensuring that its sensitivity varies approximately inversely with the background light, a relationship known as Weber’s law (Weber, 1834) and one that we discuss in detail in the next section. Because of this adaptation, the eye is more sensitive to changes in light level than to a steady input. -

1P MON. PM Cortex and Subcortical Auditory Nuclei
dynamic role for the vestibular system in orientation and flight control. Laboratory studies of flight behavior under illuminated and dark conditions in both static and rotating obstacle tests were carried out while administering heavy water ͑D2O͒ to bats to impair their vestibular inputs. Eptesicus carried out complex maneuvers through both fixed arrays of wires and a rotating obstacle array using both vision and echolocation, or when guided by echolocation alone. When treated with D2O in combination with lack of visual cues, bats showed considerable decrements in performance. These data indicate that big brown bats use both vision and echolocation to provide spatial registration for head position information generated by the vestibular system. 2:55 1pAB3. Bat’s auditory system: Corticofugal feedback and plasticity. Nobuo Suga ͑Dept. of Biol., Washington Univ., One Brookings Dr., St. Louis, MO 63130͒ The auditory system of the mustached bat consists of physiologically distinct subdivisions for processing different types of biosonar information. It was found that the corticofugal ͑descending͒ auditory system plays an important role in improving and adjusting auditory signal processing. Repetitive acoustic stimulation, cortical electrical stimulation or auditory fear conditioning evokes plastic changes of the central auditory system. The changes are based upon egocentric selection evoked by focused positive feedback associated with lateral inhibition. Focal electric stimulation of the auditory cortex evokes short-term changes in the auditory 1p MON. PM cortex and subcortical auditory nuclei. An increase in a cortical acetylcholine level during the electric stimulation changes the cortical changes from short-term to long-term. There are two types of plastic changes ͑reorganizations͒: centripetal best frequency shifts for expanded reorganization of a neural frequency map and centrifugal best frequency shifts for compressed reorganization of the map. -

Krogh's Principle
Introduction to Neuroscience: Behavioral Neuroscience Neuroethology, Comparative Neuroscience, Natural Neuroscience Nachum Ulanovsky Department of Neurobiology, Weizmann Institute of Science 2017-2018, 2nd semester Principles of Neuroethology Neuroethology seeks to understand the mechanisms by which the Neurobiology central nervous system controls the Neuroethology Ethology natural behavior of animals. • Focus on Natural behaviors: Choosing to study a well-defined and reproducible yet natural behavior (either Innate or Learned behavior) • Need to study thoroughly the animal’s behavior, including in the field: Neuroethology starts with a good understanding of Ethology. • If you study the animals in the lab, you need to keep them in conditions as natural as possible, to avoid the occurrence of unnatural behaviors. • Krogh’s principle 1 Krogh’s principle August Krogh Nobel prize 1920 “For such a large number of problems there will be some animal of choice or a few such animals on which it can be most conveniently studied. Many years ago when my teacher, Christian Bohr, was interested in the respiratory mechanism of the lung and devised the method of studying the exchange through each lung separately, he found that a certain kind of tortoise possessed a trachea dividing into the main bronchi high up in the neck, and we used to say as a laboratory joke that this animal had been created expressly for the purposes of respiration physiology. I have no doubt that there is quite a number of animals which are similarly "created" for special physiological -

Chromatic Mach Bands: Behavioral Evidence for Lateral Inhibition in Human Color Vision
Perception &: Psychophysics 1987, 41 (2), 173-178 Chromatic Mach bands: Behavioral evidence for lateral inhibition in human color vision COLIN WARE University ofNew Brunswick, Fredericton, New Brunswick, Canada and Wll..LIAM B. COWAN National Research Council of Canada, Ottawa, Ontario, Canada Apparent hue shifts consistent with lateral inhibition in color mechanisms were measured, in five purely chromatic gradients, for 7 observers. Apparent lightness shifts were measured in achro matic versions of the same gradients, and a correlation was found to exist between the magni tude of the chromatic shifts and the magnitude ofthe achromatic shifts. The data can be modeled by a function that approximates the activity of color-sensitive neurons found in the monkey cor tex. Individual differences and the failure of some previous researchers to find the effect can be accounted for by differences in the function parameters for different observers. One of the intensity gradients used by Ernst Mach an achromatic component, chromatic shifts could be (1886/1967) to demonstrate what have become known as caused by a coupling to the achromatic effect through, "Mach bands" is graphed in Figure 1 (curve C). Ob for example, the Bezold-Briicke effect. They also argue servers of a pattern with this luminance profile report a that, because prolonged viewing was allowed, chromatic bright band at the point where the luminance begins to Mach bands may be an artifact ofsuccessive contrast and decline. As with others like it, thisobservation represents eye movements and not evidence for lateral interactions. one of the principal sources ofevidence for spatially in On the other side, those who find chromatic Mach bands hibitory interactions in the human visual system. -

Seeing in Three Dimensions: the Neurophysiology of Stereopsis Gregory C
Review DeAngelis – Neurophysiology of stereopsis Seeing in three dimensions: the neurophysiology of stereopsis Gregory C. DeAngelis From the pair of 2-D images formed on the retinas, the brain is capable of synthesizing a rich 3-D representation of our visual surroundings. The horizontal separation of the two eyes gives rise to small positional differences, called binocular disparities, between corresponding features in the two retinal images. These disparities provide a powerful source of information about 3-D scene structure, and alone are sufficient for depth perception. How do visual cortical areas of the brain extract and process these small retinal disparities, and how is this information transformed into non-retinal coordinates useful for guiding action? Although neurons selective for binocular disparity have been found in several visual areas, the brain circuits that give rise to stereoscopic vision are not very well understood. I review recent electrophysiological studies that address four issues: the encoding of disparity at the first stages of binocular processing, the organization of disparity-selective neurons into topographic maps, the contributions of specific visual areas to different stereoscopic tasks, and the integration of binocular disparity and viewing-distance information to yield egocentric distance. Some of these studies combine traditional electrophysiology with psychophysical and computational approaches, and this convergence promises substantial future gains in our understanding of stereoscopic vision. We perceive our surroundings vividly in three dimen- lished the first reports of disparity-selective neurons in the sions, even though the image formed on the retina of each primary visual cortex (V1, or area 17) of anesthetized cats5,6. -

Lateral Inhibition Effects Demonstrate That the Maturational State of Neurons That Then Inhibit Their Neighbors
522 Lateral Inhibition effects demonstrate that the maturational state of neurons that then inhibit their neighbors. When a the learner’s brain is crucial for the attainment of stimulus (such as a bar of light or any other stim- a language system. ulus) excites a number of neurons in the network Several questions remain open about the mecha- (in this case neurons 4e, 5e, 6e, and 7e), the effect nisms underlying language learning. One issue is of inhibition is to suppress the neurons just out- whether specific aspects of language acquisition side the edge of the bar (3e and 8e) because those should be attributed to language-specific versus neurons are inhibited but not excited. Further, general-purpose learning mechanisms. Another issue because the neurons just inside the edges of the is whether children’s native language can affect the bar (4e and 7e) are excited by light and only inhib- way they think, and whether language is necessary ited by one neighbor, they are especially active. or helpful for the development of human concepts. This leads to perceptual contrast enhancement at borders. Further research showed that lateral inhi- Anna Papafragou bition also applied to overlapping stimuli, and that its strength fell off with distance between the See also Aphasias; Audition: Cognitive Influences; Context Effects in Perception; Speech Perception; Top- interacting stimuli. Down and Bottom-Up Processing; Word Recognition Haldan Keffer Hartline won the Nobel Prize in 1967 for discovering lateral inhibition and its neu- ral correlates. The first inhibitory circuit in the Further Readings nervous system was found in the horseshoe crab (Limulus polyphemus).