Ejemplaraccsesoabierto Arganto
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Herbarium News
Herbarium News University of Reading Herbarium Newsletter Issue No. 45, December 2008. ISSN 0953-0080 URL: www.herbarium.reading.ac.uk INTRODUCTION Welcome to another Christmas Herbarium News. As with every edition we have much to report! During the last year a number of important things have happened, not least the refurbishment of two of our taxonomy laboratories (more details below). We have also changed the name of our building from ‘Plant Science Laboratories’ to ‘The Harborne Building’ to commemorate Jeffrey Harborne, FRS, our late lamented phytochemist. This is in line with University policy to name buildings after people, and the new biomolecular sciences building (behind Chemistry) for our medical and pharmacy researchers will be called the Hopkins Building, after the late Professor Harold Hopkins, sometime Professor in Physics here who developed fibre optics and the endoscope. The Herbarium has been especially busy having now completed the incorporation of the Herbarium from the University of Southampton, again see below. HERBARIUM REVIEW AND STRATEGY Our School of Biological Sciences recently undertook a review of the Herbarium, along with the Cole Museum of Zoology and a number of other issues. Ours was led by Professor Philip John with Professor Martin Bell (Archaeology), Kate Arnold-Foster (Head of the University’s Museums and Special Collections Service and Dr Karen Henderson (Head of Technical Services, Biological Sciences). They were very positive and have recommended that a Herbarium Strategy be drawn up for the next five years, currently in progress. Senate also positively noted the Herbarium’s growing level of engagement with the University and the likely increase in the commercial value of this resource. -

Gathered Food Plants in the Mountains
34063_Rivera.qxd 7/2/07 2:03 PM Page 1 Gathered Food Plants in the Mountains of Castilla–La Mancha (Spain): Ethnobotany and Multivariate Analysis1 Diego Rivera*,2, Concepción Obón3, Cristina Inocencio3, Michael Heinrich4, Alonso Verde2, José Fajardo2, and José Antonio Palazón5 2 Departamento de Biología Vegetal, Facultad de Biología, Universidad de Murcia, 30100 Murcia, Spain 3 Departamento de Biología Aplicada, EPSO, Universidad Miguel Hernández, 03312 Orihuela, Alicante, Spain 4 Centre for Pharmacognosy and Phytotherapy, The School of Pharmacy, Univ. London, 29–39 Brunswick Sq. London, WC1N 1AX, United Kingdom 5 Departamento de Ecología e Hidrología, Facultad de Biología, Universidad de Murcia, 30100 Murcia, Spain * Corresponding author: Departamento de Biología Vegetal, Facultad de Biología, Universidad de Murcia, 30100 Murcia, Spain; e-mail: [email protected] GATHERED FOOD PLANTS IN THE MOUNTAINS OF CASTILLA–LA MANCHA (SPAIN): ETHNOBOTANY AND MULTIVARIATE ANALYSIS. Gathered food plants (GFPs) (wild and weeds) are crucial for under- standing traditional Mediterranean diets. Combining open interviews and free–listing ques- tionnaires, we identified 215 GFP items, i.e., 53 fungi and 162 from 154 vascular plant species. The variation in frequency and in salience among the items follows a rectangular hyperbola. Highly salient species were Silene vulgaris (Moench) Garcke, Scolymus hispani- cus L., and Pleurotus eryngii (DC.: Fr.) Quélet. Salience and frequency showed no correlation with the expected health benefits of each species. Regional frequency in the Mediter- ranean and local frequency are directly related. Thus, local food plants are much less “local” than expected. Different types of culinary preparations provide the most information in the cluster analysis of variables. -

Contribucion Al Estudio Palinologico De La Familia Iridaceae En Andalucia Occidental (Excepto El Genero Iris L)
Lagascalia 15(2): 189-198 (1990) CONTRIBUCION AL ESTUDIO PALINOLOGICO DE LA FAMILIA IRIDACEAE EN ANDALUCIA OCCIDENTAL (EXCEPTO EL GENERO IRIS L) M. MARTÍN CACAO & I. FERNÁNDEZ Departamento de Biología Vegetal y Ecología, Universidad de Sevilla. (Recibido el 25 de Octubre de 1988) Resumen. En este trabajo se describe la morfología del polen de 10 tazones, pertene- cientes a los géneros Gynandriris Par!., Crocus L., Romulea Maratti y Gladiolus L., exis- tentes en Andalucía Occidental, que han sido estudiados a microscopía óptica y micros- copía electrónica de barrido. En general el polen es heteropolar o apolar, bisimétrico, a veces radiosimétrico o asimétrico, tamaño mediano o grande, sistema apertural monoa- nasulcado, espiraperturado o inaperturado y téctum reticulado-perforado o equinulado- perforado. Se pone de manifiesto el caracter euripolínico del grupo y se describen cuatro tipos polínicos en base, fundamentalmente, al sistema apertura!. Summary. Pollen morphology of 10 taxa belonging to the genera Gynandriris Par!., Crocus L., Romulea Maratti and Gladiolus L., from W Andalucia are described by light and scanning electron microscopy. In general, the pollen is heteropolar or apolar, bi- symmetric, radiosymmetric or less frequently asymmetric, of large or medium siz£, 1-su!- cate, spiraperturate or nonaperturate with reticulate-perforate or equinulate-perforate tec- tum . The euripalynous condition of the group is clear and four ponen types have been distinguished, where the most variable character is the apertura! system. INTRODUCCION La familia Iridaceae cuenta con aproximadamente 1.500 especies distribuidas por todo el mundo (CRoNQuÍsT, 1981) y está representada en Andalucía Occidental (SO de España) por 24 taxones, agrupados en seis géneros incluidos en tres tribus, como reconoce VALDÉS (1987): I. -

A Detailed Study of Crocus Carpetani Series
wjpmr, 2016, 2(2), 51-53 SJIF Impact Factor: 3.535 Review Article Saxena. WORLD JOURNAL OF PHARMACEUTICAL World Journal of Pharmaceutical and Medical Research ISSN 2455-3301 AND MEDICAL RESEARCH www.wjpmr.com WJPMR A DETAILED STUDY OF CROCUS CARPETANI SERIES *Dr. R. B. Saxena Drug Standardisation Research Section, Central Research Institute- Ayurveda, Aamkho, GWALIOR- 474009 (INDIA). *Correspondence for Author: Dr. R. B. Saxena Drug Standardisation Research Section, Central Research Institute- Ayurveda, Aamkho, GWALIOR- 474009 (INDIA). Article Received on 09/01/2016 Article Revised on 01/02/2016 Article Accepted on 22/02/2016 ABSTRACT [1] [2] [3] [4] [5] [6] [7] Crocus series : C.aleppici , Autumn crocus , C. biflori , C.biflorus, C. flavi , C. longiflori , C. oriental , [8] [9] [10] [11] [12] C. sativus , C. scardici , C. speciosi , C. verni and C. versicolores have been studied. Now the sub- species Crocus-crocus carpetani series are closely related and are difficult to be separated taxonomically and have a complex cytology. Botany of crocus carpetani series, taxonomy of their species and their infra-specific taxa are presented, and their distribution, ecology and phenology, description and chromosome counts are provided with key of their identification. KEYWORDS: Crocus, Geographic area, Classification, Chromosome, Cytology, Carpetani- series, Phenology. INTRODUCTION[13-21] Such chromosome barriers are of obvious importance and can be lead on the further divergene which may The genus crocus L. consists currently of about 160 eventually give rise to acceptable species. The closely recognized species occurring from W Europe and NW related species have been difficult to separate Africa to W China, with the center of species diversity taxonomically and have also been found to be complex on the Balkan Peninsula and in Turkey. -
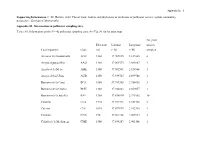
Supporting Information. C. M. Herrera. 2020. Flower Traits, Habitat, and Phylogeny As Predictors of Pollinator Service: a Plant Community Perspective
Appendix S1 – 1 Supporting Information. C. M. Herrera. 2020. Flower traits, habitat, and phylogeny as predictors of pollinator service: a plant community perspective. Ecological Monographs. Appendix S1. Information on pollinator sampling sites. TABLE S1. Information on the N = 42 pollinator sampling sites. See Fig. S1 for location map. No. plant Elevation Latitude Longitude species Local toponym Code (m) (º N) (º W) sampled Arenales del Guadalentín AGU 1360 37.909835 2.837405 4 Arroyo Aguaderillos AAG 1180 37.961573 2.883887 1 Arroyo de la Mesa AME 1100 37.902591 2.928966 1 Arroyo de los Ubios AUB 1250 37.939725 2.899766 1 Barranco de la Canal BCA 1580 37.789385 2.956438 3 Barranco de la Charca BCH 1360 37.942843 2.859057 1 Barranco de la Juanfría BJU 1380 37.836814 2.975302 10 Calarilla CLA 1710 37.944316 2.851708 2 Calerón CLE 1075 37.897973 2.942710 1 Cantalar CAN 770 37.964158 2.907974 3 Cañada de la Medianega CME 1580 37.894287 2.903146 1 Appendix S1 – 2 Cañada Pajarera CPA 1640 37.927456 2.805615 1 Collado del Calvario CCA 1420 37.950202 2.885484 1 Collado Zamora CZA 1420 37.839329 2.999615 2 Coto del Valle CVA 850 37.930542 2.928517 7 Cuesta del Bazar CBA 1330 37.906523 2.907470 1 Cuevas Bermejas CBE 1160 37.966959 2.851117 5 Fuente Bermejo FBE 1550 37.932268 2.840672 14 Fuente la Reina FRE 1460 37.941698 2.833632 2 Guadalentín GUA 1310 37.900404 2.836944 8 Hoyos de Muñoz HMU 1110 37.984831 2.876597 3 La Cabrilla LCA 1600 37.932364 2.780637 5 La Fresnedilla LFR 1080 37.980387 2.855875 2 La Mesa LME 1610 37.889823 2.913788 2 Las -

Spring Flowers of Western Andalucia
Spring Flowers of Western Andalucia Naturetrek Tour Report 21 – 28 March 2007 Spanish Festoon Narcissus cuatrecasasii Images and report compiled by Sophie Miller Naturetrek Cheriton Mill Cheriton Alresford Hampshire SO24 0NG England T: +44 (0)1962 733051 F: +44 (0)1962 736426 E: [email protected] W: www.naturetrek.co.uk Tour Report Western Andalucia Tour leader: Sophie Miller Tour participants: Kit Surrey Meg Surrey Ross Baker Lynn Whitfield Sue Bailey Chantal Pallot Alan Woods Beryl Ratcliffe Day 1 Wednesday 21st March Leaving Malaga airport we found a delightful picnic spot just off the roadside. Within a few metres of the cheese and wine there were hidden yellow anemones Anemone palmata, yellow bee orchid Ophrys lutea, Barbary nut Gyandris sisyrincium and remains of Asphodelus aestivus. After lunch a brief perusal found, typical Southern Spanish flora of Aristolochia baetica (host plant of the butterfly Spanish festoon), Cistus albidus and C. monspeliensis, Helianthemum organifolium, Turpentine tree Pistachio terebinthes, Tree germander Teucrium fruticans and the fan palm Chamaerops humilis. Up the slope a marvelous pink butterfly orchid Orchis papilionacea was found plus bee orchid Ophrys apifera and bumblebee orchid Ophrys bombyliflora. Scattered over a path was a pretty blue orchid, Ophris iricolor subsp durieui (subsp. atlantica), a plant of Southern Iberia and a very localized species. Many Spanish festoon butterflies were out enjoying the heat of the sun as much as we were. Dartford warbler, chaffinch and long-tailed tits were flying about the scrub. White clumps in the pines were noticed and housed the larvae of the pine processionary moth. A booted eagle was also spotted flying into the sun. -

Bulbous Plants in the Iberian Peninsula. the Collection of the Royal Botanical Garden Madrid
Horticulture International Journal Research Article Open Access Bulbous plants in the Iberian Peninsula. The collection of the Royal Botanical Garden Madrid Abstract Volume 4 Issue 1 - 2020 168 species of bulbous from Liliaceae, Amaryllidaceae and Iridaceae families are found Silvia Villegas, Carlos Aedo, Ramón Morales in the Iberian Peninsula and the Balearic Islands, 16 of them are introduced, numerous Real Jardín Botánico, CSIC. Plaza de Murillo 2. 28014 Madrid, endemic and subendemic (also from the other side of the Pyrenees in France). The living Spain bulb collection at the Royal Botanic Garden Madrid conserves and manages most of the species growing in the wild in the Iberian Peninsula. Correspondence: Ramón Morales, Real Jardín Botánico, CSIC. Plaza de Murillo 2. 28014 Madrid, Spain, Tel 914203017, Fax Keywords: bulbous, living collection, iberian peninsula, liliaceae, amaryllidaceae, 914200157, Email iridaceae Received: December 20, 2019 | Published: January 30, 2020 Introduction Iridaceae family have recently established themselves in the wild, like Ixia paniculata, Watsonia meriana, Freesia refracta, Freesia alba The geophites in the sense of Raunkiaer are herbaceous perennial and Crocosmia × crocosmiflora. Sisyrinchium, with 2 species living plants with its perennating buds below the ground. They can be in the Iberian Peninsula, has been introduced from North America rhizomes, tubers, bulbs and corms, these last solid bulbs almost and occurs in a few localities in Spain. All the bulbous plants species 1 without cataphylls. These types of plants are very important in the of the three families are included in Table 2. In the case of endemic Mediterranean region, because their biological cycle is well adapted (e) and subendemic (s), subspecies are also included. -

Spring Flowers of Western Andalucia
Spain - Spring Flowers of Western Andalucia Naturetrek Tour Report 13 - 20 March 2018 Ophrys scolopax Narcissus bulbocodium Narcissus cantabricus fl Report & photos by Paul Harmes Naturetrek Mingledown Barn Wolf's Lane Chawton Alton Hampshire GU34 3HJ UK T: +44 (0)1962 733051 E: [email protected] W: www.naturetrek.co.uk Tour Report Spain - Spring Flowers of Western Andalucia Tour participants: Paul Harmes (leader) and eight Naturetrek clients Day 1 Tuesday 13th March Fly London Gatwick to Malaga – Transfer to Ronda area Eight tour group members met with Paul at Gatwick Airport’s North Terminal, for the Easyjet flight, EZY8605, to Malaga. Upon our arrival, we made our way to our hire vehicle, in the multi-storey car park. We left Malaga airport in a north-westerly direction, past the cargo area, via the perimeter road towards El Peron, where we took the A7052 for several miles before joining the A357, Campilos road. We continued north-west, passing the ‘white village’ of Ardales on our left, and on to the junction with the A367, the Ronda road. Along the way, Spotless Starlings, Barn Swallows, Crested Larks, Griffon Vultures and Short-toed Snake Eagle were seen. After a light snack at the El Meson de Diego venta, we continued on the A367 south, through Cuevas del Becerro, to the outskirts of Ronda, where we joined the A376, Seville road, west for about eight kilometres to our final destination, the Hotel Cortijo Las Piletas, our accommodation for the week. On arrival, we were met by Elisenda, our host, who quickly settled us into our rooms. -

First Updated Checklist of the Vascular Flora of Andalusia (S of Spain), One of the Main Biodiversity Centres in the Mediterranean Basin
Phytotaxa 339 (1): 001–095 ISSN 1179-3155 (print edition) http://www.mapress.com/j/pt/ PHYTOTAXA Copyright © 2018 Magnolia Press Monograph ISSN 1179-3163 (online edition) https://doi.org/10.11646/phytotaxa.339.1.1 PHYTOTAXA 339 First updated checklist of the vascular flora of Andalusia (S of Spain), one of the main biodiversity centres in the Mediterranean Basin MIGUEL CUETO1,*, MANUEL MELENDO2, ESTHER GIMÉNEZ1, JULIÁN FUENTES3, ENRIQUE LÓPEZ CARRIQUE4 & GABRIEL BLANCA5 1 Departamento de Biología y Geología, CECOUAL, Universidad de Almería, Crta. Sacramento s/n, La Cañada de S. Urbano, ES- 04120 Almería, Spain; e-mail: [email protected], [email protected] 2 Departamento de Biología Animal, Biología Vegetal y Ecología. Facultad de Ciencias Experimentales, Universidad de Jaén, Campus Las Lagunillas s/n, ES-23071 Jaén, Spain; e-mail: [email protected] 3 C/ Castillo 5, bajo F, ES-18140 La Zubia, Granada, Spain; e-mail: [email protected] 4 Departamento de Educación, CECOUAL, Universidad de Almería, Crta. Sacramento s/n, La Cañada de S. Urbano, ES-04120 Almería, Spain; e-mail: [email protected] 5 Departamento de Botánica, Facultad de Ciencias, Universidad de Granada, C/ Fuentenueva s/n, ES-18001 Granada, Spain; e-mail: [email protected] *author for correspondence Magnolia Press Auckland, New Zealand Accepted by Manuel B. Crespo: 24 Dec. 2017; published: 20 Feb. 2018 MIGUEL CUETO, MANUEL MELENDO, ESTHER GIMÉNEZ, JULIÁN FUENTES, ENRIQUE LÓPEZ CARRIQUE & GABRIEL BLANCA First updated checklist of the vascular flora of Andalusia (S of Spain), one of the main biodiversity centres in the Mediterranean Basin (Phytotaxa 339) 95 pp.; 30 cm. -
Ème Contribution À I'qtube Mr,5 Elante.« Tomatique Et Jlébttinaie Bu »Art Aaatioual Me Zlemten Axonomie, Qtotoie, Qatactéttttque Que
République Algérienne Démocratique et Populaire Ministère de l'Enseignement Supérieur et de la Recherche Scientifique UNIVERSITE ABOU BEKR BELKAID TLEMCEN Faculté des Sciences de la Nature et de la Vie, des Sciences de la Terre et de l'Univers Département d'Ecologie et Environnement Laboratoire de recherche Promotion des Ressources Hydriques, Pédologiques et Minières THESE Me Présentée par: FELIDJ Menel En vue de l'obtention du Diplôme de Doctorat En Ecologie et Environnement Option : Ecologie végétale .... ème Contribution à I'Qtube Mr,5 elante.« tomatique et JlébttinaIe bu »art Aaatioual Me Zlemten axonomie, Qtotoie, Qatactéttttque que ., Soutenue le: 15/09/ 2011, devant le jury composé de: Directeur de Thèse : M. BOUAZZA Mohamed. Professeur Université ABOU BEKR BELKAID, TLEMCEN Président: M. BENSALAR Mustapha. Professeur Université AB OU BEKR BELKAID, TLEMCEN Examinateurs: M. BENABADJT Noury. Professeur Université ABOU BEKR BELKAID, TLEMCEN M. IIELLEL Benchabane. Professeur Université Djilali LYABES, SIPI BEL ABBES M. MEHDADI Zoheir. Professeur Université Djilali LYABES, SIDI BEL ABBES M. HASNAOUI Oukacha. MCC Université de SAIDA Année universitaire 2010/2011 4 L fl L J tJ7 JJJ fl Li j L:A ) Jl Ld & J) & L4 (:Pi 1 tA J L LJi LA ç 4)i iJ • L 4 J 4 JI , 2 J LI Résumé: L'Algérie, de par la diversité de son climat (méditerranéen, aride) et de ses sols, possède une flore particulièrement riche en plantes médicinales et aromatiques, dont la plupart existe à l'état spontané. L'inventaire et la valorisation de ces plantes médicinales et aromatiques demeure un domaine de grande importance pour le pays car c'est une source de produits à haute valeur ajoutée. -

Flora Vascular De Andalucía Oriental
Flora Vascular de Andalucía Oriental Flora Vascular de Andalucía Oriental Editores científicos: Gabriel Blanca (Universidad de Granada) Baltasar Cabezudo (Universidad de Málaga) Miguel Cueto (Universidad de Almería) Carlos Fernández López (Universidad de Jaén) Concepción Morales Torres (Universidad de Granada) Volumen 1 SELAGINELLACEAE–CERATOPHYLLACEAE FLORA VASCULAR DE ANDALUCÍA ORIENTAL Edita: Consejería de Medio Ambiente. Junta de Andalucía Consejera de Medio Ambiente Cinta Castillo Jiménez Viceconsejero de Medio Ambiente Juan Jesús Jiménez Martín Director General de Gestión del Medio Natural Francisco Javier Rojo Madrid Dirección Facultativa Rosa María Mendoza Castellón Diseño Gráfico, Maquetación y Retoque fotográfico Miguel Carrión Publicidad S.L El Ejido (Almería) Fotomecánica e impresión Escandón Impresores. Sevilla © 2009, Texto y fotografías: los autores © 2009, Consejería de Medio Ambiente Quedan rigurosamente prohibidas, sin la autorización escrita de los titulares del “Copyright” y bajo las sanciones establecidas en las leyes, la reproducción total o parcial de esta obra por cualquier medio o procedimiento, comprendidos la reprografía y el tratamiento informático, y la distribución de ella mediante venta o alquiler. ISBN obra completa: 978–84–92807–12–3 ISBN volumen 1: 978–84–92807–13–0 Depósito legal: SE-4430-2009 (I) Impreso en España 2009 La obra completa debe citarse como: Blanca G., Cabezudo B., Cueto M., Fernández López C. & Morales Torres C. (2009, eds.). Flora Vascular de Andalucía Oriental, 4 vols. Consejería de Medio Ambiente, Junta de Andalucía, Sevilla. Este libro debe citarse como: Blanca G., Cabezudo B., Cueto M., Fernández López C. & Morales Torres C. (2009, eds.). Flora Vascular de Andalucía Oriental. Volumen 1: Selaginellaceae–Ceratophyllaceae. Consejería de Medio Ambiente, Junta de Andalucía, Sevilla. -

An Updated Checklist of the Vascular Flora of Sierra Nevada (SE Spain)
Phytotaxa 261 (1): 001–057 ISSN 1179-3155 (print edition) http://www.mapress.com/j/pt/ PHYTOTAXA Copyright © 2016 Magnolia Press Article ISSN 1179-3163 (online edition) http://dx.doi.org/10.11646/phytotaxa.261.1.1 An updated checklist of the vascular flora of Sierra Nevada (SE Spain) JUAN LORITE Dpto. de Botánica, Universidad de Granada, ES-18071 Granada, Spain; e-mail: [email protected] Abstract To have an updated checklist available for a given area is crucial for many purposes (educational, conservation, manage- ment, etc.). In this paper, a complete and updated checklist of the vascular flora of Sierra Nevada mountain range (SE Spain), a recognized hotspot for plant diversity in the Mediterranean basin, is presented. It includes 2,353 taxa, 359 more than in the previous checklist (an increase of 15.25%). Also, data are shown on composition (in terms of families, number of hybrids, native/alien species, endemics, etc.) and threat status according to the IUCN. The result is a complete and updated checklist flora of this recognised biodiversity Mediterranean hotspot. Keywords: Baetic Mountains, Endemics, Iberian Peninsula, Plant Biodiversity Hotspot, Threatened flora, Vascular Plants Introduction The floristic inventory of a given area is the very first basis, a prerequisite, and a starting point for assessing plant conservation, management, and ecological restoration, providing information on the need for additional surveys or data collections, as well as establishing the starting point for more detailed studies (Kier et al. 2005). It aids in identifying and correctly naming species, essential resources for biodiversity estimates and biogeographic studies. Furthermore, an accurate checklist of species fulfils important social functions, since the richness value is often the simplest and the most direct way of presenting the concept of biodiversity to the society in general.