Novel Insights Into Sabino1 and Splashed White Coat Color Patterns in Horses
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Absolutely Arabians, LLC (Caledonia, WI)
Absolutely Arabians, LLC (Caledonia, WI) Sport Horse Disciplines: Dressage In-hand Performance Horses for Sale? Yes Stallion(s) At Stud? Yes Youngstock For Sale? Yes Absolutely Arabians, LLC is a small Arabian horse breeding farm located in southeastern Wisconsin. Our goal is to produce beautiful and athletic Breeding Stock For Sale? Arabians and Half Arabians through quality breeding. Sometimes The main focus of our breeding program is our beautiful Multi National Training Offered? Champion Black Arabian Stallion Tuxedo Thyme ABA++++// . Tux has four AHA National Championships in Sport Horse In-Hand and FEI Level No Dressage proving he has great conformation and is very athletic. He has also won the USDF All Breeds Award for FEI Level Dressage. In total Tux Contact: has won 16 AHA National Titles and 44 AHA Regional Titles in Arabian Halter, Sport Horse In Hand, Sport Horse Show Hack and Dressage. Barb Suvaka 4030 West 6 1/2 Mile Road In addition to Tux, we also have several mares that we are focusing on Caledonia, WI 53108 breeding for Sport Horse disciplines. One is a full sister to Tux named (262) 880-1943 Essence of Thyme ABA. The other is new to us and more info will be published on her in 2018. [email protected] Website: http://www.tuxedothyme.com Additional Link: www.facebook.com/tuxedothyme Velvet Tuxedo 2015 Arabian filly (Tuxedo Thyme ABA x Sun-Kist Mekisha Tuxedo Thyme ABA++++// 2004 stallion (Pyro Thyme SA x SMA Alad Khelima) Photos © Suzanne Sturgill &© Fire and Earth Photography An Tearmann Arabians (Lodi, WI) Sport Horse Disciplines: Dressage In-hand SHUS and/or SHSH Performance Horses for Sale? Yes An Tearmann Arabians is located in the rolling hills of south central Wisconsin Stallion(s) At Stud? near Lodi. -

O'bajan Senior (1881-1910) the Majestic Black Stallion 1910 ~ A
American Shagya Arabian Verband Newsletter Holiday Issue Merry Christmas and Happy New Year 2010 The Majestic Black Stallion 1910 ~ A One Hundred Year Legacy ~ 2010 by Ildiko Mohammed-Ziegler and Shasa Gornic, Dr. med. vet. rendering from a photograph dated 1909 O’Bajan Senior (1881-1910) Page 2 American Shagya Arabian Verband Newsletter To the reader This splendid article was originally written in the Hungarian language and then translated into English. As English is not the primary language of the authors, parts of the translation lacked clarity. It was not our intention to Anglicize the article but to enhance the translation into a more readable document. Some of the original sentences remain a bit awkward, but in their context they are understandable. Please also note that the much of the text quoted in italics was taken from books and docu ments whose original language was also not English, but Hungarian and German. Many of these quotations are from texts written in the late 1800's and are not easily understandable as to our current use of the language. However we did not feel that we should alter the text from their original sources. O'Bajan, an original Arabian (purebred ox) the desert a horse to fall in love with.” He great stud farms will emphasize the im was born in 1880 in TellelKelach, Syria. continues...”O'Bajan is small, only 154 cm, pression.”(5) His sire was O'Bajan senior 18811910 but of him, one may say there is a ‘multum and his dam was Maneghie. in pravo’ (much in little). -

Identification of Copy Number Variants in Horses
Downloaded from genome.cshlp.org on October 1, 2021 - Published by Cold Spring Harbor Laboratory Press Research Identification of copy number variants in horses Ryan Doan,1 Noah Cohen,2 Jessica Harrington,2 Kylee Veazy,2 Rytis Juras,3 Gus Cothran,3 Molly E. McCue,4 Loren Skow,3 and Scott V. Dindot1,5,6 1Department of Veterinary Pathobiology, 2Department of Large Animal Clinical Sciences, 3Department of Veterinary Integrative Biosciences, College of Veterinary Medicine and Biomedical Sciences, Texas A&M University, College Station, Texas 77843, USA; 4Department of Veterinary Population Medicine, University of Minnesota College of Veterinary Medicine, St. Paul, Minnesota 55108, USA; 5Department of Molecular and Cellular Medicine, Texas A&M Health Science Center College of Medicine, College Station, Texas 77843, USA Copy number variants (CNVs) represent a substantial source of genetic variation in mammals. However, the occurrence of CNVs in horses and their subsequent impact on phenotypic variation is unknown. We performed a study to identify CNVs in 16 horses representing 15 distinct breeds (Equus caballus) and an individual gray donkey (Equus asinus) using a whole- exome tiling array and the array comparative genomic hybridization methodology. We identified 2368 CNVs ranging in size from 197 bp to 3.5 Mb. Merging identical CNVs from each animal yielded 775 CNV regions (CNVRs), involving 1707 protein- and RNA-coding genes. The number of CNVs per animal ranged from 55 to 347, with median and mean sizes of CNVs of 5.3 kb and 99.4 kb, respectively. Approximately 6% of the genes investigated were affected by a CNV. Biological process enrichment analysis indicated CNVs primarily affected genes involved in sensory perception, signal transduction, and metabolism. -

List of Horse Breeds 1 List of Horse Breeds
List of horse breeds 1 List of horse breeds This page is a list of horse and pony breeds, and also includes terms used to describe types of horse that are not breeds but are commonly mistaken for breeds. While there is no scientifically accepted definition of the term "breed,"[1] a breed is defined generally as having distinct true-breeding characteristics over a number of generations; its members may be called "purebred". In most cases, bloodlines of horse breeds are recorded with a breed registry. However, in horses, the concept is somewhat flexible, as open stud books are created for developing horse breeds that are not yet fully true-breeding. Registries also are considered the authority as to whether a given breed is listed as Light or saddle horse breeds a "horse" or a "pony". There are also a number of "color breed", sport horse, and gaited horse registries for horses with various phenotypes or other traits, which admit any animal fitting a given set of physical characteristics, even if there is little or no evidence of the trait being a true-breeding characteristic. Other recording entities or specialty organizations may recognize horses from multiple breeds, thus, for the purposes of this article, such animals are classified as a "type" rather than a "breed". The breeds and types listed here are those that already have a Wikipedia article. For a more extensive list, see the List of all horse breeds in DAD-IS. Heavy or draft horse breeds For additional information, see horse breed, horse breeding and the individual articles listed below. -

Special Issue Horse Genetics DIRECTOR’S Message
CENTER FOR EQUINE HEALTH SCHOOL OF VETERINARY MEDICINE • UNIVERSITY OF CALIFORNIA, DAVIS SUMMER 2020 Special Issue Horse Genetics DIRECTOR’S Message s an equine genetics researcher, I am particularly excited to share A this special issue of the Horse Report with you. Inside, you will find a roadmap to many of the currently available equine genetic tests, including the AQHA “five-panel” test, and more. The equine genome sequence was published in 2009, the result of a years- long collaborative effort by the international equine research community. This resource drastically changed how researchers approach equine genetics and accelerated the rate of discovery. Increased availability and affordability allowed the application of advanced molecular tools to equine diseases and traits. As a result, genetic tests are available in a variety of breeds. Most available tests are for simple, Mendelian diseases and traits – those caused by a single gene or locus. Complex diseases and traits likely involve more than one gene and may be influenced by environmental effects. The 2018 release of a new equine genome sequence assembly, coupled with cost reductions that make whole-genome sequencing possible for large numbers of horses, are enabling research in these areas. As an equine geneticist and veterinarian, I am especially interested in applying whole genome sequencing and advanced diagnostic tools to equine precision medicine. This highly individualized approach will focus on early detection and prevention of disease, taking into account both genetic information and environmental factors. The idea is to target individuals based on their clinical condition as well as their unique body chemistry and genetics. -

Electronic Supplementary Material - Appendices
1 Electronic Supplementary Material - Appendices 2 Appendix 1. Full breed list, listed alphabetically. Breeds searched (* denotes those identified with inherited disorders) # Breed # Breed # Breed # Breed 1 Ab Abyssinian 31 BF Black Forest 61 Dul Dülmen Pony 91 HP Highland Pony* 2 Ak Akhal Teke 32 Boe Boer 62 DD Dutch Draft 92 Hok Hokkaido 3 Al Albanian 33 Bre Breton* 63 DW Dutch Warmblood 93 Hol Holsteiner* 4 Alt Altai 34 Buc Buckskin 64 EB East Bulgarian 94 Huc Hucul 5 ACD American Cream Draft 35 Bud Budyonny 65 Egy Egyptian 95 HW Hungarian Warmblood 6 ACW American Creme and White 36 By Byelorussian Harness 66 EP Eriskay Pony 96 Ice Icelandic* 7 AWP American Walking Pony 37 Cam Camargue* 67 EN Estonian Native 97 Io Iomud 8 And Andalusian* 38 Camp Campolina 68 ExP Exmoor Pony 98 ID Irish Draught 9 Anv Andravida 39 Can Canadian 69 Fae Faeroes Pony 99 Jin Jinzhou 10 A-K Anglo-Kabarda 40 Car Carthusian 70 Fa Falabella* 100 Jut Jutland 11 Ap Appaloosa* 41 Cas Caspian 71 FP Fell Pony* 101 Kab Kabarda 12 Arp Araappaloosa 42 Cay Cayuse 72 Fin Finnhorse* 102 Kar Karabair 13 A Arabian / Arab* 43 Ch Cheju 73 Fl Fleuve 103 Kara Karabakh 14 Ard Ardennes 44 CC Chilean Corralero 74 Fo Fouta 104 Kaz Kazakh 15 AC Argentine Criollo 45 CP Chincoteague Pony 75 Fr Frederiksborg 105 KPB Kerry Bog Pony 16 Ast Asturian 46 CB Cleveland Bay 76 Fb Freiberger* 106 KM Kiger Mustang 17 AB Australian Brumby 47 Cly Clydesdale* 77 FS French Saddlebred 107 KP Kirdi Pony 18 ASH Australian Stock Horse 48 CN Cob Normand* 78 FT French Trotter 108 KF Kisber Felver 19 Az Azteca -

Lipizzan Laurels United States Lipizzan Federation©
Lipizzan Laurels United States Lipizzan Federation© Awards Program The United States Lipizzan Federation (USLF) recognizes Lipizzans/XL Lipizzans competing in events against all breeds with two special award programs—the USLF Lipizzan/XL Lipizzan Laurels Award and the USLF Lipizzan/XL Lipizzan Star Award. Award participants automatically are entered in both programs when they submit results. The USLF Lipizzan Laurels presents a Lipizzan Laurels Award for outstanding performance by a Lipizzan/XL Lipizzan horse in several sections within the program’s ten major disciplines. Junior Exhibitor Awards are also presented in all ten disciplines. Lipizzan Laurels awards are tabulated on an annual basis during the competition year, which runs from November 1st of the previous year to October 31st of the current year. The Lipizzan/XL Lipizzan Star Award is a lifetime award presented to each Lipizzan/XL Lipizzan meeting Star requirements. Lipizzan/XL Lipizzan horses may take as many years as needed to earn Star points. The owner of the achieving horse will receive an official bronze, silver, gold, or platinum Star when they have accumulated the necessary points in their discipline. Stars are presented in ten disciplines: Show, Competitive Trail, Endurance, Dressage, Eventing, Working Western, Carriage Pleasure, Combined Driving, Western Dressage, and Working Equitation. Horses who earn five of the ten bronze Stars are presented with the USLF Lipizzan/XL Lipizzan Sport Horse Award. 1 GENERAL GUIDELINES To be eligible for these awards, horses must be registered with the USLF at the time scores are earned. Both the owner and all riders or drivers of the horse must be current USLF members at the time scores are earned. -

Quantitative Genetic Analysis of Melanoma and Grey Level in Lipizzan Horses
7th World Congress on Genetics Applied to Livestock Production, August 19-23, 2002, Montpellier, France QUANTITATIVE GENETIC ANALYSIS OF MELANOMA AND GREY LEVEL IN LIPIZZAN HORSES I. Curik1, M. Seltenhammer2 and J. Sölkner3 1Animal Science Department, Faculty of Agriculture, University of Zagreb, Croatia 2Clinic of Surgery and Ophthalmology, University of Veterinary Medicine Vienna, Austria 3Department of Livestock Science, University of Agricultural Sciences Vienna, Austria INTRODUCTION Changes of coat colour in which the "dark (non-grey)" colour present in foals is progressively replaced by grey is a known phenomenon in horses. A similar process is present in humans. The grey coat colour is inherited as a dominant trait and is the characteristic, although not exclusive, colour for some horse breeds (Bowling, 2000). The Lipizzan horse, originally bred for show and parade at the Imperial Court in Vienna, is among those breeds. Unfortunately, melanomas (skin tumours) are more prevalent in grey than in non-grey horses (e.g. Seltenhammer, 2000). The causative relationship for this positive association as well as the molecular basis for both traits (melanoma and grey level) are not known (Rieder, 1999 ; Seltenhammer, 2000). The inheritance of coat colour in horses has been always studied from a qualitative view (Sponenberg, 1996 ; Bowling, 2000). In the present study we quantified the grey level (shade) and estimated the proportion of additive genetic component (heritability in the narrow sense) of this trait. Further, we estimated the genetic relationship between melanoma stages and grey level as well as the additive inheritance of melanoma stages. MATERIAL AND METHODS Horses. Data for this study was collected from 351 grey Lipizzan horses of four national studs (Djakovo – Croatia ; 64 horses, Piber – Austria ; 160 horses, Szilvesvarad – Hungary ; 67 horses and Topol'cianky – Slovakia ; 60 horses). -
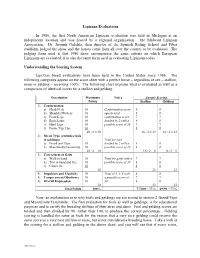
2-Evaluation Summary (H0933573
Lipizzan Evaluations In 1986, the first North American Lipizzan evaluation was held in Michigan at an independent location and was hosted by a regional organization – the Midwest Lipizzan Association. Dr. Jaromir Oulehla, then director of the Spanish Riding School and Piber studfarm, judged the show and the horses came from all over the country to be evaluated. The judging form used at that 1986 show encompasses the same criteria on which European Lipizzans are evaluated; it is also the same form used in evaluating Lipizzans today. Understanding the Scoring System Lipizzan breed evaluations have been held in the United States since 1986. The following categories appear on the score sheet with a perfect horse – regardless of sex – stallion, mare or gelding – receiving 100%. The following chart explains what is evaluated as well as a comparison of identical scores for a stallion and gelding. Description Maximum Notes Sample Scoring Points Stallion Gelding 1) Conformation a) Head/Neck 10 Conformation score 8 8 b) Shoulder/Withers 10 equals total 8 8 c) Front Legs 10 confirmation score 7 7 d) Back/Loins 10 divided by 2 with a 8 8 e) Hind Legs 10 possible score of 30 7 7 f) Frame/Top Line 10 8 8 60 / 2 = 30 46 / 2 = 23 46 / 2 = 23 2) Breed Type (excludes foals & geldings) Total for type a) Breed and Type 10 divided by 2 with a 8 0 b) Masculinity/Femininity 10 possible score of 10 8 0 20 / 2 = 10 16 / 2 = 8 0 / 2 = 0 3) Correctness of Gaits a) Walk in hand 10 Total for gaits with a 7 7 b) Trot in hand and free 10 possible score of 30 8 8 c) Canter free 10 8 8 30 23 23 4) Impulsion and Elasticity 10 Total of 4, 5, 6 with 8 8 5) Temperament/Obedience 10 a possible score of 7 7 6) Overall Impression 10 30 8 8 30 23 23 Total Points 100% 77/100 = 77% 69/90 = 77% Now, an explanation as to why foals and geldings are not scored in section 2 Breed/Type and Masculinity/Femininity. -

Arabian Coat Color Patterns
Arabian Coat Color Patterns Copyright 2011 Brenda Wahler In the Arabian breed, there are three unusual coat colors or patterns that occur in some purebred horses. The first is sabino, the only white spotting pattern seen in purebred Arabians, characterized by bold white face and leg markings, and, in some cases, body spotting. The second pattern is rabicano, a roan-like intermixture of white and dark hairs. Both sabino and rabicano horses are often registered by their base coat color, with white patterns noted as markings, but some extensively marked individuals have been registered as “roan,” even though true roan is a separate coat color. The third unusual coat color is dominant white, a mutation characterized by a predominantly white hair coat and pink skin, present at birth. All Arabians in the United States currently known to be dominant white trace to a single stallion, foaled in 1996, verified to be the offspring of his registered Arabian parents, both of whom were solid-colored. It is difficult to know how many Arabians have these unusual colors as they are often not searchable in registration records. For many years, Arabians with dominant white, body spots, or simply “too much white” were discouraged from registration, and white body markings were penalized in halter classes. The exclusion of boldly-marked “cropout” horses was also common in other registries, leading to the formation of a number of color breed associations. However, when parentage verification became possible, horses born with “too much” white could be confirmed as the offspring of their stated parents, and breed registries generally relaxed their rules or policies that previously excluded such animals. -

Impact of White Spotting Alleles, Including W20, on Phenotype in the American Paint Horse
bioRxiv preprint doi: https://doi.org/10.1101/678052; this version posted June 21, 2019. The copyright holder for this preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. Running Head: White spotting in the American Paint Horse Title: Impact of white spotting alleles, including W20, on phenotype in the American Paint Horse Samantha A. Brooks*, Katelyn M. Palermo*, Alisha Kahn*, and Jessica Hein# *University of Florida Department of Animal Sciences, UF Genetics Institute, Gainesville FL, 32611-0910 #American Paint Horse Association, Fort Worth TX, 76161-0023 Acknowledgments: The authors would like to thank the many APHA staff members for their efforts in submitting and collating the data analyzed in this study. Thanks to the UF undergraduate researchers who generously volunteered for data-entry work on this project: Hannah Hillard, Kalisse Horne, Rachel Kullman, Erica Riano, Matt Winter, Courtney McCreary, Rachel Shepherd, Anna Moskovitz, and Kaycie Miller. Our gratitude to Dr. Ernie Bailey for proofreading the manuscript. bioRxiv preprint doi: https://doi.org/10.1101/678052; this version posted June 21, 2019. The copyright holder for this preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. Abstract: The American Paint Horse Association (APHA) officially records pedigree and performance information for their breed; these registered stock-type horses are valued for utility in work on the farm and ranch and as pleasure horses. As the name of the breed implies, the breed is also valued for attractive white spotting patterns on the coat. -

The Effect of the Size and Shape of Chestnuts on the Identification of Horses
ACTA UNIVERSITATIS AGRICULTURAE ET SILVICULTURAE MENDELIANAE BRUNENSIS Volume LX 3 Number 6, 2012 THE EFFECT OF THE SIZE AND SHAPE OF CHESTNUTS ON THE IDENTIFICATION OF HORSES I. Bihuncová, E. Sobotková, T. Petlachová, M. Píšová, M. Kosťuková, H. Černohorská Received: July 10, 2012 Abstract BIHUNCOVÁ, I., SOBOTKOVÁ, E., PETLACHOVÁ, T., PÍŠOVÁ, M., KOSŤUKOVÁ, M., ČERNOHORSKÁ, H.: The eff ect of the size and shape of chestnuts on the identifi cation of horses. Acta univ. agric. et silvic. Mendel. Brun., 2012, LX, No. 6, pp. 21–32 A study was carried out on 11 horse breeds comprising of the Akhal Teke (n = 23); English Thoroughbred (n = 23); Arabian Thoroughbred (n = 18); Czech Warm-Blood (n = 21); Old Kladrubian horse (n = 20); Hucul horse (n = 20); Czech – Moravian Belgian horse (n = 20); Noriker horse (n = 7); Silesian Noriker (n = 14); Hafl inger (n = 20); Shetland pony (n = 20) to determine the shape and size of chestnuts. Chestnuts of 206 horses classifi ed in three phylogeny classes were measured and drawn. The necessary data (breed; sex; name; sire; dam; sire of dam; age of horse; colour of horse; colour of the chestnut; bone) were entered into a special form. In the form the outlines of the shapes of the chestnuts were drawn; using a calliper we measured the protrusion of the chestnut at its highest point and the width at the widest part of the chestnut. We found no identical or similar shapes of the chestnuts within the breed or phylogeny class. We confi rmed that the outlines of the chestnuts can be used as identifying marks because they are unique for each horse.