Composition of Eggs of Several Bird Species
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Incubation Constancy in the Red-Winged Blackbird
INCUBATION CONSTANCY IN THE RED-WINGED BLACKBIRD LARRY C. HOLCOMB Avian incubation behavior is affected by a multitude of exogenous and en- dogenous factors. Kendeigh (1952, 196313) and Skutch (1962) reviewed in- cubation in many different orders of birds and discussed factors affecting the amount of time spent in incubation. Among workers recently reporting on incubation behavior in wild passerines are Prescott (1964)) Mumford (1964)) Erpino (1968)) Maxwell and Putnam (1972)) and Morton et al. (1972). In the Red-winged Blackbird (Ag e1 aius phoeniceus) , Nero (1956a, 195613) has published observations on female behavior during the repro- ductive cycle, but nothing was reported on the incubation constancy (per- cent of daylight hours spent on the nest). I have reported (Holcomb, 1968, 1970) that female Redwings incubated normal-sized artificial eggs a mean of 19.4 days before abandoning them. This is 8.4 days beyond the normal incubation period. The present study was designed to determine the incubation constancy in the egg-laying period, to discover if it increased each day as the incubation behavior developed, and to find if there was less incubation each day as females neared the day when eggs were abandoned in prolonged incubation. METHODS I studied the incubation behavior of Red-winged Blackbirds near Omaha, Nebraska, in 1968 and 1969. Birds were breeding in a variety of habitats, including weed, alfalfa, and clover fields, hedgerows, ditch banks, and marshes. I visited the nesting areas nearly every day, beginning in March and ending in August. Male Redwings generally arrived in early March and females soon afterward. -

Incubating and Hatching Eggs
EPS-001 7/13 Incubating and Hatching Eggs Gregory S. Archer and A. Lee Cartwright* hether eggs come from a common chicken Factors that affect hatchability or an exotic bird, you must store and incu- W Breeder Hatchery bate them carefully for a successful hatch. Envi- Breeder nutrition Sanitation ronmental conditions, handling, sanitation, and Disease Egg storage record keeping are all important factors when it Mating activity Egg damage comes to incubating and hatching eggs. Egg damage Incubation—Management of Correct male and female setters and hatchers Fertile egg quality body weight Chick handling A fertile egg is alive; each egg contains living cells Egg sanitation that can become a viable embryo and then a chick. Egg storage Eggs are fragile and a successful hatch begins with undamaged eggs that are fresh, clean, and fertile. Collecting and storing fertile eggs You can produce fertile eggs yourself or obtain Fertile eggs must be collected carefully and stored them elsewhere. While commercial hatcheries properly until they are incubated. Keeping the produce quality eggs that are highly fertile, many eggs at proper storage temperatures keeps the do not ship small quantities. If you mail order embryo from starting and stopping development, eggs, be sure to pick them up promptly from your which increases embryo mortality. Collecting receiving area. Hatchability will decrease if eggs eggs frequently and storing them properly delays are handled poorly or get too hot or too cold in embryo development until you are ready to incu- transit. bate them. If you produce the eggs on site, you must care for the breeding stock properly to ensure maximum Egg storage reminders fertility. -

Care of Fertile Eggs Prior to Incubation 1
CARE OF FERTILE EGGS PRIOR TO INCUBATION 1. Keep eggs at 50º - 60º F (room temperature). (DON’T PUT IN A REFRIGERATOR, IT IS TOO COLD!) 2. Store the eggs with the BIG end up in egg cartons. 3. You can store for 10 days after they have been laid before hatch rate decreases (50% hatch rate is a good rate). Incubation / hatching time begins once the eggs are placed in the incubator and brought up to 99 ½ºF. Count day 1 after first 24 hours. Incubation/hatching time for chickens are 21 days and 28 to 33 days for ducks. INCUBATOR SET UP 1. Place the incubator(s) away from any windows as the sunlight will magnify thru the plexi-glass cover making it too hot. 2. Set up the incubator four (4) hours prior to adding eggs; it will give the incubator an opportunity to Diagram #2 Round Corner regulate the water temperature in the reservoir. Diagram #1 Put water into both of the troughs on the square Square corner incubator incubator cornered incubators (see diagram #1). In the round corner model which has a larger and smaller trough, place water in the outside trough labeled “circulating” (see diagram #2). (Be sure that the incubator turns on at 99º and off at 100ºF.) 3. Place the plastic thermometer, simply lay it across the top of the eggs, (see diagram #5). It will basically ride on top of the eggs. 4. Cover the adjusting stem on top with paper cup taped to help eliminate the likelihood of the setting being accidentally changed. -

Mercury and Selenium Contamination in Waterbird Eggs and Risk to Avian Reproduction at Great Salt Lake, Utah
Prepared in cooperation with the U.S. Fish and Wildlife Service Mercury and Selenium Contamination in Waterbird Eggs and Risk to Avian Reproduction at Great Salt Lake, Utah Open-File Report 2015–1020 U.S. Department of the Interior U.S. Geological Survey Cover: Franklin’s gulls nesting at Bear River Migratory Bird Refuge, Great Salt Lake, Utah. Photograph taken by Josh Ackerman in 2012. Mercury and Selenium Contamination in Waterbird Eggs and Risk to Avian Reproduction at Great Salt Lake, Utah By Joshua T. Ackerman, Mark P. Herzog, C. Alex Hartman, John Isanhart, Garth Herring, Sharon Vaughn, John F. Cavitt, Collin A. Eagles-Smith, Howard Browers, Chris Cline, and Josh Vest Prepared in cooperation with the U.S. Fish and Wildlife Service Open-File Report 2015–1020 U.S. Department of the Interior U.S. Geological Survey U.S. Department of the Interior SALLY JEWELL, Secretary U.S. Geological Survey Suzette M. Kimball, Acting Director U.S. Geological Survey, Reston, Virginia: 2015 For more information on the USGS—the Federal source for science about the Earth, its natural and living resources, natural hazards, and the environment—visit http://www.usgs.gov or call 1–888–ASK–USGS For an overview of USGS information products, including maps, imagery, and publications, visit http://www.usgs.gov/pubprod To order this and other USGS information products, visit http://store.usgs.gov Any use of trade, firm, or product names is for descriptive purposes only and does not imply endorsement by the U.S. Government. Although this information product, for the most part, is in the public domain, it also may contain copyrighted materials as noted in the text. -

Embryonic Exposure to Oestrogen Causes Eggshell Thinning and Altered Shell Gland Carbonic Anhydrase Expression in the Domestic Hen
REPRODUCTIONRESEARCH Embryonic exposure to oestrogen causes eggshell thinning and altered shell gland carbonic anhydrase expression in the domestic hen C Berg, A Blomqvist1, L Holm1, I Brandt, B Brunstro¨m and Y Ridderstra˚le1 Department of Environmental Toxicology, Uppsala University, Norbyva¨gen 18 A, 753 36 Uppsala, Sweden and 1Department of Anatomy and Physiology, Swedish University of Agricultural Sciences, Box 7045, 750 07 Uppsala, Sweden Correspondence should be addressed to C Berg; Email: [email protected] Abstract Eggshell thinning among wild birds has been an environmental concern for almost half a century. Although the mechanisms for contaminant-induced eggshell thinning are not fully understood, it is generally conceived to originate from exposure of the laying adult female. Here we show that eggshell thinning in the domestic hen is induced by embryonic exposure to the syn- thetic oestrogen ethynyloestradiol. Previously we reported that exposure of quail embryos to ethynyloestradiol caused histo- logical changes and disrupted localization of carbonic anhydrase in the shell gland in the adult birds, implying a functional disturbance in the shell gland. The objective of this study was to examine whether in ovo exposure to ethynyloestradiol can affect eggshell formation and quality in the domestic hen. When examined at 32 weeks of age, hens exposed to ethynyloestra- diol in ovo (20 ng/g egg) produced eggs with thinner eggshells and reduced strength (measured as resistance to deformation) compared with the controls. These changes remained 14 weeks later, confirming a persistent lesion. Ethynyloestradiol also caused a decrease in the number of shell gland capillaries and in the frequency of shell gland capillaries with carbonic anhy- drase activity. -

Class Three: Breeding
Class Three: Breeding WHERE DO SEABIRDS LAY THEIR EGGS? Nest or no nest? • Some species use no nesting material. E.g., the white tern lays a single egg on an open branch. • Some species use a very little bit of nesting material. E.g., the tufted puffin may use a few pieces of grass and a couple of feathers. • Some species build more substantial nests. E.g. kittiwakes cement their nests onto small cliff shelves by trampling mud and guano to form a base. And, some cormorants build large nests in trees from sticks and twigs. [email protected] www.seabirdyouth.org 1 White tern • Also called fairy tern. • Tropical seabird species. • Lays egg on branch or fork in tree. No nest. • Newly hatched chicks have well developed feet to hang onto the nesting-site. White Tern. © Pillot, via Creative Commons. On the coast or inland? • Most seabird species breed on the coast and offshore islands. • Some species breed fairly far inland, but still commute to the ocean to feed. E.g., kittlitz’s murrelets nest on scree slopes on coastal mountains, and parents may travel more than 70km to their feeding grounds. • Other species breed far inland and never travel to the ocean. E.g., double crested cormorants breed on the coast, but also on lakes in many states such as Minnesota. [email protected] www.seabirdyouth.org 2 NESTING HABITAT (1) Ground Some species breed on the ground. These species tend to breed in areas with little or no predation, such as offshore islands (e.g., terns and gulls) or in the Antarctic (e.g., penguins, albatross). -

Egg Laying in Pet Birds Egg Laying in Pet Birds Can Be a Serious Health Threat
Egg Laying in Pet Birds Egg laying in pet birds can be a serious health threat. This article explains why (and what to do if) your bird starts laying eggs. Providing proper, non-incandescent lighting, a healthy diet, and adequate sleep, as well as removing nesting toys or materials are key to discouraging egg laying. Overview In wild birds and breeding birds, egg laying is a natural, seasonal process. However, female pet birds can also lay eggs, even without the presence of a male. Such eggs are infertile and will not hatch, even if incubated. A bird in the peak of health on an ideal diet may be able to sustain some egg production without serious harm. However, with captive pet birds, it can also become an obsession, because the eggs do not hatch and allow the full cycle to complete, thus turning off the hormonal trigger to lay eggs. Constant egg laying will deplete your bird of vital nutrients, and predispose her to malnutrition , osteoporosis, and life-threatening health problems, such as egg binding and yolk peritonitis. While egg laying can occur in any breed, it is most common in cockatiels, lovebirds, budgies, canaries, and finches. Egg laying can start anytime from 5 months to over 10 years of age. If you find an egg, you want to immediately correct any environmental factors that predispose your bird to lay eggs. If that does not work, your bird may require medical treatment to control egg laying, so you’ll want to get her to a qualified Avian vet. There are several safe, effective hormonal treatments available, which your Avian vet can tailor to your bird's needs. -

Consumption of Bird Eggs by Invasive
WWW.IRCF.ORG/REPTILESANDAMPHIBIANSJOURNALTABLE OF CONTENTS IRCF REPTILES IRCF& AMPHIBIANS REPTILES • VOL &15, AMPHIBIANS NO 4 • DEC 2008 • 19(1):64–66189 • MARCH 2012 IRCF REPTILES & AMPHIBIANS CONSERVATION AND NATURAL HISTORY TABLE OF CONTENTS INTRODUCED SPECIES FEATURE ARTICLES . Chasing Bullsnakes (Pituophis catenifer sayi) in Wisconsin: On the Road to Understanding the Ecology and Conservation of the Midwest’s Giant Serpent ...................... Joshua M. Kapfer 190 . The Shared HistoryConsumption of Treeboas (Corallus grenadensis) and Humans on ofGrenada: Bird Eggs A Hypothetical Excursion ............................................................................................................................Robert W. Henderson 198 byRESEARCH Invasive ARTICLES Burmese Pythons in Florida . The Texas Horned Lizard in Central and Western Texas ....................... Emily Henry, Jason Brewer, Krista Mougey, and Gad Perry 204 1 2 3 . The Knight Anole (Anolis equestrisCarla) inJ. Florida Dove , Robert N. Reed , and Ray W. Snow .............................................Brian J. Camposano, Kenneth L. Krysko, Kevin M. Enge, Ellen M. Donlan, and Michael Granatosky 212 1 SmithsonianCONSERVATION Institution, Division ALERT of Birds, NHB E-600, MRC 116, Washington, District of Columbia 20560, USA ([email protected]) 2U.S. Geological Survey, Fort Collins Science Center, 2150 Centre Ave, Bldg C, Fort Collins, Colorado 80526, USA ([email protected]) . World’s Mammals in Crisis ............................................................................................................................................................ -

Nest Building and Egg Laying by Redwinged Blackbirds in Response to Artificial Manipulations
NEST BUILDING AND EGG LAYING BY REDWINGED BLACKBIRDS IN RESPONSE TO ARTIFICIAL MANIPULATIONS LARRY C. HOLCOMB THE responsesof breedingbirds to many external stimuli have been well reviewedby Lehrman (1959), Eisner (1960), and van Tienhoven (1961). The studiesperformed by Emlen (1941) on the Tricolored Blackbird (Agelaiustricolor) are especiallyrelevant to this paper because of somesimilarities of this icterid to the RedwingedBlackbird (Agelaius phoeniceus).Orians (1961) reportsthe similaritiesand differencesin the ecologyand social systemsof the Redwingedand Tricolored Blackbirds. The objectivesof the presentstudy were to assesssome of the external stimuli that affect physiologicaland related behavioral patterns in the Red-wing. The two nmin objectiveswere to 1) determineresponses of Red- wings to artificial nest manipulations,compared with thosereported by Emlen (1941) for the TricoloredBlackbird, and 2) discoverif Red-wings are determinateor indeterminatelayers. Emlen found that the TricoloredBlackbird usually lays its first egg on the day followingnest completion.Payne (1965) reportsthat Tricolored Blackbirdsusually lay their first eggin the morningfollowing completion of the nestbut that Red-wingfemales have a rangeof 1-4 days. In addition Payne notes that all Tricolored Blackbirds had ovulated on the day of lining the nest, whereassix of eight Red-wing femalestaken from lined nestshad not ovulated. I have made similar observationson Red-wings. Payne statesthat "clutchsize of RedwingedBlackbirds is apparentlydeter- mined by the developmentof the ovary a few days before the final egg is laid." Holcomb and Twiest (1968) present data on the time between nest completionand egglaying for 42 uplandand marshbreeding Red-wings in May and June. Theseshow a meandelay in laying of 2.0 days after nest completion,with a range of between0 and 5 days. -

Hatching Small Numbers of Eggs
PNW 478 Reprinted October 1995 Hatching small numbers of eggs J.C. Hermes ncubation is the process by which the embryo within the egg develops into a fully formed chick capable of breaking free from the shell. Incubat- Iing eggs requires several important environmental considerations if any chicks are to hatch. Of primary concern are temperature and relative humidity. Attention to secondary considerations can result in increased numbers of eggs hatching. Natural vs. artificial relative humidity within the proper limits to hatch chicks. incubation Artificial incubation is an The first decision to be made excellent alternative to the is whether to incubate eggs broody hen. naturally or artificially. Natural incubation uses a broody hen to Temperature incubate eggs by sitting on them Correct incubation tempera- in a nest. Broody hens, when ture depends on the type of available, work best for small incubator. In forced draft incuba- clutches of eggs. tors, those in which a fan is Broodiness, the behavior of a present, temperature should be incubators, humidity is attained setting hen, has been bred out of ° maintained between 99.5 F and by filling a pan or some other some chickens. Others don’t care ° 100 F for most bird eggs. In still reservoir in the incubator with for chicks effectively. Some air incubators, where no fan breeds commonly used to hatch water. is present, a temperature of For most eggs, a relative eggs include New Hampshires, between 101°F and 103°F Plymouth Rocks, Rhode Island humidity of between 55 and measured at the top of the egg 60 percent is adequate during the Reds, and Cochins. -

Common Birds of the Estero Bay Area
Common Birds of the Estero Bay Area Jeremy Beaulieu Lisa Andreano Michael Walgren Introduction The following is a guide to the common birds of the Estero Bay Area. Brief descriptions are provided as well as active months and status listings. Photos are primarily courtesy of Greg Smith. Species are arranged by family according to the Sibley Guide to Birds (2000). Gaviidae Red-throated Loon Gavia stellata Occurrence: Common Active Months: November-April Federal Status: None State/Audubon Status: None Description: A small loon seldom seen far from salt water. In the non-breeding season they have a grey face and red throat. They have a long slender dark bill and white speckling on their dark back. Information: These birds are winter residents to the Central Coast. Wintering Red- throated Loons can gather in large numbers in Morro Bay if food is abundant. They are common on salt water of all depths but frequently forage in shallow bays and estuaries rather than far out at sea. Because their legs are located so far back, loons have difficulty walking on land and are rarely found far from water. Most loons must paddle furiously across the surface of the water before becoming airborne, but these small loons can practically spring directly into the air from land, a useful ability on its artic tundra breeding grounds. Pacific Loon Gavia pacifica Occurrence: Common Active Months: November-April Federal Status: None State/Audubon Status: None Description: The Pacific Loon has a shorter neck than the Red-throated Loon. The bill is very straight and the head is very smoothly rounded. -
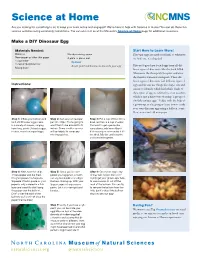
Make a DIY Dinosaur Egg (PDF)
Science at Home Are you looking for something to do to keep your brain active and engaged? We’re here to help with Science at Home! You can do these fun science activities using commonly found items. You can also visit us at the Museum’s Science at Home page for additional resources. Make a DIY Dinosaur Egg Materials Needed: Start Here to Learn More! Balloons Wooden mixing spoon Dinosaur eggs are rarely fossilized, so whenever Newspaper or other thin paper A plate or place-mat we find one, it’s a big deal. 1 cup water Optional 1 cup all-purpose flour Acrylic paint and brushes to decorate your egg Paleontologists have found eggs from all dif- Mixing bowl ferent types of dinosaurs, like the duck-billed Maiasaura, the theropod Oviraptor, and even the massive titanosaur sauropods. These dif- ferent types of dinosaurs laid different types of Instructions: eggs and we can use things like shape, size and texture to identify which laid which. Each of these types of egg are referred to as an ootaxon, which is just a fancy way of saying “a group of similarly unique eggs.” Today, with the help of a grown-up, you’re going to learn how to make your own dinosaur eggs using a balloon, some flour, water and old newspaper. Step 1: Inflate your balloon and Step 2: Cut up your newspa- Step 3: Put a cup of flour into a tie it off. Dinosaur eggs came per into strips. You’re going to bowl, and mix in a cup of water.