Composition Patterns of Ornamental Flora in the Czech Republic
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Foliar Anatomy of the Genus Silene L. (Caryophyllaceae) at Sectional Level in Iran
DOI: http://dx.doi.org/10.22092/ijb.2016.107954 FOLIAR ANATOMY OF THE GENUS SILENE L. (CARYOPHYLLACEAE) AT SECTIONAL LEVEL IN IRAN M. Nejati Edalatian, F. Ghahremaninejad & F. Attar Received 2015. 06. 08; accepted for publication 2016. 11. 02 Nejati Edalatian, M., Ghahremaninejad, F. & Attar, F. 2016. 12. 30: Foliar anatomy of the genus Silene L. (Caryophyllaceae) at sectional level in Iran. -Iran. J. Bot. 22 (2): 138-158. Tehran. In this research, Iranian species of 20 sections of the genus Silene L. have been anatomically studied for the first time. Some of the most important anatomical characters of leaf in this genus are listed here: general shape of leaves in transverse section, type of stomata cells, form of trichome, type of mesophyll (dorsiventral or isobilateral), presence or absence and type of collenchyma, thickness of sclerenchyma, existence or lack of water-storage, shape of vascular bundles, number of parenchyma layers in midrib, form and number of spongy and palisade parenchyma, presence or absence of parenchyma bundle sheath, number of vascular bundles in midrib. Based on our results we are able to identify and classify sections of this genus (tables 2, 3). In addition, we selected the most important characters related to midrib and lamina. All of these results confirm anatomical characters have taxonomic value for separating sections in the genus Silene. Maliheh Nejati Edalatian & Farrokh Ghahremaninejad (correspondence<[email protected]>) Department of Plant Sciences, Faculty of Biological Sciences, Kharazmi University, 43 Dr. Mofatteh Avenue, Postal code 15719-14911, Tehran, Iran. -Farideh Attar, Tehran University, College of Sciences, School of Biology, Department of Botany, Central Herbarium, Tehran, Iran. -

Journal of the Royal Horticultural Society of London
I 3 2044 105 172"381 : JOURNAL OF THE llopl lortimltoal fbck EDITED BY Key. GEORGE HEXSLOW, ALA., E.L.S., F.G.S. rtanical Demonstrator, and Secretary to the Scientific Committee of the Royal Horticultural Society. VOLUME VI Gray Herbarium Harvard University LOXD N II. WEEDE & Co., PRINTERS, BEOMPTON. ' 1 8 8 0. HARVARD UNIVERSITY HERBARIUM. THE GIFT 0F f 4a Ziiau7- m 3 2044 i"05 172 38" J O U E N A L OF THE EDITED BY Eev. GEOEGE HENSLOW, M.A., F.L.S., F.G.S. Botanical Demonstrator, and Secretary to the Scientific Committee of the Royal Horticultural Society. YOLUME "VI. LONDON: H. WEEDE & Co., PRINTERS, BROMPTON, 1 8 80, OOUITOIL OF THE ROYAL HORTICULTURAL SOCIETY. 1 8 8 0. Patron. HER MAJESTY THE QUEEN. President. The Eight Honourable Lord Aberdare. Vice- Presidents. Lord Alfred S. Churchill. Arthur Grote, Esq., F.L.S. Sir Trevor Lawrence, Bt., M.P. H. J". Elwes, Esq. Treasurer. Henry "W ebb, Esq., Secretary. Eobert Hogg, Esq., LL.D., F.L.S. Members of Council. G. T. Clarke, Esq. W. Haughton, Esq. Colonel R. Tretor Clarke. Major F. Mason. The Rev. H. Harpur Crewe. Sir Henry Scudamore J. Denny, Esq., M.D. Stanhope, Bart. Sir Charles "W. Strickland, Bart. Auditors. R. A. Aspinall, Esq. John Lee, Esq. James F. West, Esq. Assistant Secretary. Samuel Jennings, Esq., F.L S. Chief Clerk J. Douglas Dick. Bankers. London and County Bank, High Street, Kensington, W. Garden Superintendent. A. F. Barron. iv ROYAL HORTICULTURAL SOCIETY. SCIENTIFIC COMMITTEE, 1880. Chairman. Sir Joseph Dalton Hooker, K.C.S.I., M.D., C.B.,F.R.S., V.P.L.S., Royal Gardens, Kew. -
Fire-Resistant Plants for Oregon Home Landscapes
FFire-ire-RResistantesistant PlantsPlants forfor OregonOregon HomeHome LandscapesLandscapes Suggesting specific types of vegetation that may reduce your risk from wildfire. Stephen Fitzgerald Area Extension Forester and Associate Professor Amy Jo Waldo Area Extension Horticulture Agent and Assistant Professor OSU Extension Service 1421 S. Hwy 97, Redmond, OR 97756 Introduction Oregon has many wildfire prone areas. In these places, fires are a natural part of the changing landscape. As homes are built in these areas, special precautions must be taken by the homeowner to pro- tect their property. Installation of fire- resistive roofing is critical to preventing firebrands from igniting the home from a roof fire. Well maintained fire-resistant vegetation and irrigated landscape is also critical within close proximity of a home. These actions DO NOT insure that your home will survive a wildfire, but they provide for a good chance of structural survival. Implementation of FireFree [www.firefree.org] and FireWise [www.firewise.org] activities can also significantly improve chances of a home surviving a wildfire. Fire-resistant vegetation. When landscaping around a home, most homeowners are interested in creating a landscape that is aesthetically pleasing, compliments their home, and has varia- tions in color, texture, flowers, and foliage. If your home is located in or adjacent to forests or rangeland, you should also consider the flammability of plants within your home landscape. Flammable plant material in your land- scape can increase the fire-risk around your home. The 1991 Oakland Hills Fire in California is a prime example of how flam- mable plant material (Eucalyptus trees) can act as fuel and contribute to the inten- sity of a wildfire. -

15. CARYOPTERIS Bunge, Pl. Mongholico-Chin. 27. 1835. 莸属 You Shu Cardioteucris C
Flora of China 17: 43–47. 1994. 15. CARYOPTERIS Bunge, Pl. Mongholico-Chin. 27. 1835. 莸属 you shu Cardioteucris C. Y. Wu. Herbs, subshrubs, or shrubs, erect or climbing. Leaves opposite, simple, entire or dentate, usually with glistening glands. Flowers in lax or dense cymes often aggregate into thyrses, rarely solitary. Calyx (4- or) 5- (or 6)-dentate or -lobed. Corolla short tubed, slightly 2-lipped, margin entire or dentate; lobes 5, spreading; lower lobe larger, concave, fringed. Stamens 4, often strongly exserted, inserted on apical part of corolla tube. Ovary 4-locular; ovules pendulous or laterally attached. Stigma 2-cleft. Fruit dry, usually dividing into four nutlets. Sixteen species: C and E Asia, 14 species in China. 1a. Inflorescences terminal, narrowly cylindric thyrses, sharply differentiated from vegetative shoots, cymes subtended by very reduced bracts. 2a. Thyrses lax; lateral cymes to 5-flowered; calyx clearly 2-lipped; corolla white .............................................. 12. C. siccanea 2b. Thyrses very dense; lateral cymes more than 5-flowered; calyx deeply 6-lobed; corolla bluish purple ......... 13. C. bicolor 1b. Inflorescences axillary or subterminal, mostly cymes, sometimes compact thyrses or reduced to a solitary flower, all except sometimes the uppermost subtended by normal leaves. 3a. Inflorescences dense axillary thyrses; fruit globose, ca. 4 × as long as calyx ................................................ 14. C. paniculata 3b. Inflorescences axillary or subterminal cymes, often corymbose, sometimes reduced to 1 flower; fruit usually winged or lobed, to 2 × as long as calyx. 4a. Leaf blade entire or with occasional teeth. 5a. Leaf blade linear-lanceolate or linear-oblong to narrowly oblanceolate, abaxial veins often darker than rest of leaf surface. -

(Dr. Sc. Nat.) Vorgelegt Der Mathematisch-Naturwissenschaftl
Zurich Open Repository and Archive University of Zurich Main Library Strickhofstrasse 39 CH-8057 Zurich www.zora.uzh.ch Year: 2012 Flowers, sex, and diversity: Reproductive-ecological and macro-evolutionary aspects of floral variation in the Primrose family, Primulaceae de Vos, Jurriaan Michiel Posted at the Zurich Open Repository and Archive, University of Zurich ZORA URL: https://doi.org/10.5167/uzh-88785 Dissertation Originally published at: de Vos, Jurriaan Michiel. Flowers, sex, and diversity: Reproductive-ecological and macro-evolutionary aspects of floral variation in the Primrose family, Primulaceae. 2012, University of Zurich, Facultyof Science. FLOWERS, SEX, AND DIVERSITY. REPRODUCTIVE-ECOLOGICAL AND MACRO-EVOLUTIONARY ASPECTS OF FLORAL VARIATION IN THE PRIMROSE FAMILY, PRIMULACEAE Dissertation zur Erlangung der naturwissenschaftlichen Doktorwürde (Dr. sc. nat.) vorgelegt der Mathematisch-naturwissenschaftliche Fakultät der Universität Zürich von Jurriaan Michiel de Vos aus den Niederlanden Promotionskomitee Prof. Dr. Elena Conti (Vorsitz) Prof. Dr. Antony B. Wilson Dr. Colin E. Hughes Zürich, 2013 !!"#$"#%! "#$%&$%'! (! )*'+,,&$-+''*$.! /! '0$#1'2'! 3! "4+1%&5!26!!"#"$%&'(#)$*+,-)(*#! 77! "4+1%&5!226!-*#)$%.)(#!'&*#!/'%#+'.0*$)/)"$1'(12%-).'*3'0")"$*.)4&4'*#' "5*&,)(*#%$4'+(5"$.(3(-%)(*#'$%)".'(#'+%$6(#7.'2$(1$*.".! 89! "4+1%&5!2226!.1%&&'%#+',!&48'%'9,%#)()%)(5":'-*12%$%)(5"'"5%&,%)(*#'*3' )0"';."&3(#!'.4#+$*1"<'(#'0")"$*.)4&*,.'%#+'0*1*.)4&*,.'2$(1$*.".! 93! "4+1%&5!2:6!$"2$*+,-)(5"'(12&(-%)(*#.'*3'0"$=*!%14'(#'0*1*.)4&*,.' 2$(1$*.".>'5%$(%)(*#'+,$(#!'%#)0".(.'%#+'$"2$*+,-)(5"'%..,$%#-"'(#' %&2(#"'"#5($*#1"#).! 7;7! "4+1%&5!:6!204&*!"#")(-'%#%&4.(.'*3'!"#$%&''."-)(*#'!"#$%&''$"5"%&.' $%12%#)'#*#/1*#*204&4'%1*#!'1*2$0*&*!(-%&&4'+(.)(#-)'.2"-(".! 773! "4+1%&5!:26!-*#-&,+(#!'$"1%$=.! 7<(! +"=$#>?&@.&,&$%'! 7<9! "*552"*?*,!:2%+&! 7<3! !!"#$$%&'#""!&(! Es ist ein zentrales Ziel in der Evolutionsbiologie, die Muster der Vielfalt und die Prozesse, die sie erzeugen, zu verstehen. -

American Rock Garden Society Bulletin
American Rock Garden Society Bulletin FARRERIANA — Elizabeth Hall 1 CLIFFSIDE OF FLOWERS — Fred Lape 12 PRIMULA DEORUM -Heinz Weber 13 THE AQUILEGIAS — T. J. Cole 15 AEQUAM SERVARE MENTEM — Paul H. Boswell 20 UPRIGHT FORM OF LOISELEURIA PROCUMBENS - James Baggett 22 SOME OF ISRAEL'S NATIVE PLANTS — Ruth Benjamin 23 BOOK REVIEW — Bernard Harkness 31 REQUESTS BY MEMBERS 32 CONFERENCE AT HARROGATE — Dr. Henry Tod 33 OMNIUM-GATHERUM 35 Vol. 29 JANUARY, 1971 No. 1 DIRECTORATE BULLETIN Editor Emeritus DR. EDGAR T. WHERRY, University of Pennsylvania, Philadelphia 4, Pa. Editor ALBERT M. SUTTON 9608 26th Ave. N.W., Seattle, Washington 98107 AMERICAN ROCK GARDEN SOCIETY President Emeritus HAROLD EPSTEIN, 5 Forest Court, Larchmont, New York President— BERNARD E. HARKNESS, Box 264, R.D. #1, Pre-emption Rd., Geneva, N. Y. 14456 Secretary _ RICHARD W. REDFIELD, BOX 26, Closter, N.J. 07624 Treasurer _. _ ALEX D. REID, 260 Boulevard, Mountain Lakes, N. J. Vice-Presidents BRIAN O. MULLIGAN DONALD E. HAVENS BOYD KLINE HARRY BUTLER MRS. ARMEN GEVJAN Directors Term Expires 1971 Mrs. Herbert Brinckerhoff H. Lincoln Foster Lee Raden Term Expires 1972 Mrs. Sallie D. Allen Jerome A. Lukins Henry R. Fuller Term Expires 1973 F. Owen Pearce Mrs. L. N. Roberson George Pride Director of Seed Exchange MR. HENRY R. FULLER P. O. Box 158, Easton, Connecticut 06425 Director of Slide Collection ELMER C. BALDWIN 400 Tecumseh Road, Syracuse, N.Y. 13224 REGIONAL CHAIRMEN Northwestern CLIFFORD G. LEWIS, 4725 119th Avenue S.E., Bellevue, Wash. 98004 Western F. O. PEARCE, 54 Charles Hill Road, Orinda, Calif. 94563 Midwestern MRS. -

Number 3, Spring 1998 Director’S Letter
Planning and planting for a better world Friends of the JC Raulston Arboretum Newsletter Number 3, Spring 1998 Director’s Letter Spring greetings from the JC Raulston Arboretum! This garden- ing season is in full swing, and the Arboretum is the place to be. Emergence is the word! Flowers and foliage are emerging every- where. We had a magnificent late winter and early spring. The Cornus mas ‘Spring Glow’ located in the paradise garden was exquisite this year. The bright yellow flowers are bright and persistent, and the Students from a Wake Tech Community College Photography Class find exfoliating bark and attractive habit plenty to photograph on a February day in the Arboretum. make it a winner. It’s no wonder that JC was so excited about this done soon. Make sure you check of themselves than is expected to seedling selection from the field out many of the special gardens in keep things moving forward. I, for nursery. We are looking to propa- the Arboretum. Our volunteer one, am thankful for each and every gate numerous plants this spring in curators are busy planting and one of them. hopes of getting it into the trade. preparing those gardens for The magnolias were looking another season. Many thanks to all Lastly, when you visit the garden I fantastic until we had three days in our volunteers who work so very would challenge you to find the a row of temperatures in the low hard in the garden. It shows! Euscaphis japonicus. We had a twenties. There was plenty of Another reminder — from April to beautiful seven-foot specimen tree damage to open flowers, but the October, on Sunday’s at 2:00 p.m. -

RHS Members' Seed Scheme: Seed List 2018
RHS Members’ Seed Scheme 2018 rhs.org.uk/seedlist 1 RHS Seed Collections 2018 See page 5 for collection details AGM Collection Cottage Garden Collection Chelsea Collection Shade Collection Greening Grey Britain Plants for Pollinators 2 RHS Members’ Seed Scheme The RHS Members’ Seed Scheme draws upon Orders should be made online at the Society’s diverse plant collections and rhs.org.uk/seedlist, anytime between wealth of expertise to offer members the 1 November and 31 March. Alternatively, you exclusive opportunity to buy seed harvested can request an order form and a printed copy from RHS gardens. Our seedlist is produced of our seedlist by contacting our Membership each year by a small, dedicated team of staff Services Team: and volunteers, based at Wisley, who collect, Tel: 020 3176 5810 clean and pack seed for members. Email: [email protected] The RHS aims to enrich everyone’s life through Or in writing to: plants so we hope you will be inspired to have a go at growing from seed, which can be fun and Membership Services Team (seeds) rewarding. Covering 200 species, our seedlist The Royal Horticultural Society contains a range of plants including annuals, 80 Vincent Square herbaceous perennials, trees and shrubs, some London SW1P 2PE of which are rare and unusual. If you are not ordering online and are wanting Applying for Seed to process your order by post, then you will need to place your order before the end of This year we have increased the allocation of January as paper order forms will not be sent seed packets from 12 to 15 (please note only out after this time. -

Het Geslacht Viburnum (Deel I) Een Grote Familie Met Meer Dan 860 Soorten
Viburnum x burkwoodii 'Mohawk'. Het geslacht Viburnum (deel I) Een grote familie met meer dan 860 soorten Binnen de systematische indeling van het Regnum Vegetabile – het Plantenrijk – behoort het geslacht Viburnum tot de orde van de Dipsacales en daar- binnen tot de familie van de Caprifoliaceae*, de kamperfoeliefamilie. Auteur: Jan P. Mauritz VRT De Caprifoliaceae is een grote familie met meer dan van de Caprifoliaceae is de nodige onrust onder verschillen. Zo zijn er dwergen en heel grote mak- 860 soorten, verdeeld over 42 geslachten, die voor- ‘mijn vrienden’. Er zijn verschillende lezingen en kers, tot wel 7 tot 8 meter hoog, meestal met koe- namelijk voorkomen op het noordelijk halfrond. Er andere indelingen in omloop, waarbij het geslacht pelvormige kronen. In 1908 heeft de grootmeester zijn er toch weer een paar tegen de draad in: die ondergebracht wordt in een nauw verwante fami- Alfred Rehder het geslacht ingedeeld in secties. In komen aan de andere kant van de evenaar voor, lie, Adoxaceae, of als aparte monotypische familie de jaren daarna werden er nog een aantal secties o.a. in het Andesgebergte in Zuid-Amerika. Het zijn Viburnaceae aangemerkt wordt. van gevonden en species aan toegevoegd, waar- voornamelijk klimplanten, lianen, struikvormers en door het aantal secties opgelopen is tot elf stuks. kleine boomvormers. Het natuurlijk verspreidings- Zoals in een eerder deel van het feuilleton al aan- Uit de namen van deze secties blijkt vaak of een gebied van Viburnum is enorm en omvat de gehele gegeven, zal uw schrijver er geen woorden meer species enige bekendheid geniet. Maar ook vaak aarde. -

Caryophyllaceae)
BIBL., INST. SYST. BOT., UPPSALA. Kapsel: Nordic Journal of Botany Nummer: Correction - By mistake, a draft version of this paper was published in Nord. J. Bot. 20: 513-518. The correct version is published here. A revised generic classification ofthe tribe Sileneae (Caryophyllaceae) B. Oxelman, M. Lidén, R. K. Rabeler and M. Popp Oxelman, B, Lidén, M., Rabeler, R. K. &. Popp, M. 2001. A revised generic classification of the tribe Sileneae (Caryophyllaceae) - Nord. J. Bot. 20: 743-748. Copenhagen. ISSN-0105-055X. A reclassification of the tribe Sileneae compatible with molecular data is presented. The genus Eudianthe (E. laeta and E. coeli-rosa) is restored. Viscaria, Ixoca (heliosperma), and Atocion together form a well supported monophyletic group distinct from Silene and Lychnis, and are recognized at generic level. With Agrostemma and Petrocoptis, the number of genera in the tribe sums up to eight. The new combinations Silene samojedora, Silene ajanensis, Lychnis abyssinica, Atocion asterias, Atocion compacta, Atocion lerchenfeldiana, and Atocion rupestris are made. B. Oxelman, Evolutionsbiologlskt Centrum (EBC), Uppsala Universitet, Norbyvägen 18D, SE-752 36 Uppsala, Sweden. E-mail: [email protected]. - M. Lidén,, Botaniska trädgården, Uppsala universitet, Villavägen 6, SE-752 36 Uppsala, Sweden. E-mail: [email protected]. - R. K. Rabeler, University of Michigan Herbarium, 1205 North University Ave., Ann Arbor MI 48109-1057 USA. E-mail: [email protected]. - M. Popp, Evolutionsbiologlskt Centrum (EBC), Uppsala Universitet, Norbyvägen 18D, SE-752 36 Uppsala, Sweden. E-mail: magnus. popp@ebc. uu.se. Introduction Apocynaceae (Sennblad & Bremer 1996). Careful analyses of molecular and/or morphological data have With the recent advances in biotechnology, in particular in all these eases revealed that at least one other taxon, the rapid development of the polymerase chain reaetion traditionally recognized at the same rank, is actually an (PCR) and DNA sequencing, our understanding of the ingroup in the respective taxon (i.e. -
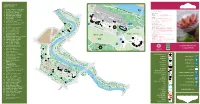
The-Commonwealth-Collection.Pdf
Round Border COMMONWEALTH Fern COLLECTION Garden Gate River W 1. Protea cyanoides - South Africa alk 2. Strelitzia reginae - South Africa GENERAL INFORMATION 3. Acacia karroo - Zambia Long Pit Glasshouse OPENING HOURS 4. Banksia ericifolia - Australia 32 GROUNDS 7 am to dusk (all year) 5. Wollemia nobilis - Australia 33 31 GLASSHOUSES 10 am - 6 pm (4.15 pm in winter) 30 6. Leptospermum scoparium - CHARGES 29 stairs New Zealand Hopkirk Glasgow City Council maintains a policy of free entry Building 7. Cyathea arborea - Jamaica 28 THE ENQUIRIES OFFICE: is situated behind the main range 27 25 15 and further information may be obtained there. 11 Enquiries 8. Ficus carica - Malta 26 24 23 20 21 22 10 19 Office PHONE: 0141 276 1614 9. Phormium tenax - New Zealand 17 Main Range FAX: 0141 276 1615 18 Glasshouses Curator’s 10. Angraecum podochiloides - 16 14 EMAIL: [email protected] House WEB: glasgowbotanicgardens.com or 13 Private Cameroon 12 www.glasgow.gov.uk/parks 11. Ansellia africana - Zambia 9 GROUPS VISITS: are especially welcome and a guide may be 12. Bowiea volubilis - Kenya Botanic Gardens available if arranged in advance with the Gardens’ Office. 13. Cycas seemannii - Fiji Tea Room DOGS: are allowed in the grounds, but should be kept on a 14. Ficus benjamina - India short leash. Dogs are not permitted in the glasshouses with the Peter exception of guide dogs. 15. Heliconia psittacorum - Walker TRANSPORT: by bus from the City Centre nos. 6, 6a, 6b, Trinidad & Tobago Memorial QUEEN 8, 8a, 10a, and 19. 16. Elettaria cardamomum - Sri Lanka MARGARET By Underground - to Hillhead Station 17. -

Lamiaceae), a Doubtful Species from Korea
Phytotaxa 195 (1): 079–085 ISSN 1179-3155 (print edition) www.mapress.com/phytotaxa/ PHYTOTAXA Copyright © 2015 Magnolia Press Article ISSN 1179-3163 (online edition) http://dx.doi.org/10.11646/phytotaxa.195.1.6 Identity of Microtoena coreana (Lamiaceae), a doubtful species from Korea QIANG WANG State Key Laboratory of Systematic & Evolutionary Botany, Institute of Botany, Chinese Academy of Sciences, Xiangshan, Beijing 00093, China. E-mail: [email protected] Abstract The species Microtoena coreana has long been extremely doubtful. This species is endemic to South Korea, and only the holotype has been collected. The author, who described this species as new, placed it into the genus Microtoena without confidence. Some taxonomists even considered that it might belong to another genus instead of Microtoena. However, no conclusion has been achieved. In the present study, the holotype of Microtoena coreana was found and carefully examined. This holotype possesses 5-lobed corolla, broadly campanulate calyxes, and long-exserted stamens, which make it distinct from all the other species of Microtoena. The results of examination on a large number of specimens and statistical analysis of multiple characters clearly show that the holotype of Microtoena coreana is an element of Tripora divaricata (= Caryop- teris divaricata). Therefore, the name Microtoena coreana is here excluded from the genus Microtoena, and is reduce to a synonym of Tripora divaricata. In addition, the lectotype of Tripora divaricata is designated in the present article. Keyword: Caryopteris, lectotypification, principal coordinate analysis, taxonomy, Tripora Introduction The genus Microtoena Prain (1889: pl. 1872) of Lamiaceae is distributed in southwestern China and adjacent regions.