The Bite Force of the Largest Fossil Rodent (Hystricognathi, Caviomorpha, Dinomyidae)
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Redalyc.A Distinctive New Cloud-Forest Rodent (Hystriocognathi: Echimyidae) from the Manu Biosphere Reserve, Peru
Mastozoología Neotropical ISSN: 0327-9383 [email protected] Sociedad Argentina para el Estudio de los Mamíferos Argentina Patterson, Bruce D.; Velazco, Paul M. A distinctive new cloud-forest rodent (Hystriocognathi: Echimyidae) from the Manu Biosphere Reserve, Peru Mastozoología Neotropical, vol. 13, núm. 2, julio-diciembre, 2006, pp. 175-191 Sociedad Argentina para el Estudio de los Mamíferos Tucumán, Argentina Available in: http://www.redalyc.org/articulo.oa?id=45713202 How to cite Complete issue Scientific Information System More information about this article Network of Scientific Journals from Latin America, the Caribbean, Spain and Portugal Journal's homepage in redalyc.org Non-profit academic project, developed under the open access initiative Mastozoología Neotropical, 13(2):175-191, Mendoza, 2006 ISSN 0327-9383 ©SAREM, 2006 Versión on-line ISSN 1666-0536 www.cricyt.edu.ar/mn.htm A DISTINCTIVE NEW CLOUD-FOREST RODENT (HYSTRICOGNATHI: ECHIMYIDAE) FROM THE MANU BIOSPHERE RESERVE, PERU Bruce D. Patterson1 and Paul M. Velazco1, 2 1 Department of Zoology, Field Museum of Natural History, 1400 S. Lake Shore Dr, Chicago IL 60605-2496 USA. 2 Department of Biological Sciences, University of Illinois at Chicago, 845 W. Taylor St, Chicago IL 60607 USA ABSTRACT: Recent surveys in Peru’s Manu National Park and Biosphere Reserve uncovered a new species of hystricognath rodent, a spiny rat (Echimyidae) with dense, soft fur. Inhabiting Andean cloud-forests at 1900 m, the new rodent belongs to a radiation of “brush- tailed tree rats” previously known only from the Amazon, Orinoco, and other lowland river drainages. Phylogenetic analysis of morphology (cranial and dental characters) unambiguously allies the new species with species of Isothrix. -

Classification of Mammals 61
© Jones & Bartlett Learning, LLC © Jones & Bartlett Learning, LLC NOT FORCHAPTER SALE OR DISTRIBUTION NOT FOR SALE OR DISTRIBUTION Classification © Jones & Bartlett Learning, LLC © Jones & Bartlett Learning, LLC 4 NOT FORof SALE MammalsOR DISTRIBUTION NOT FOR SALE OR DISTRIBUTION © Jones & Bartlett Learning, LLC © Jones & Bartlett Learning, LLC NOT FOR SALE OR DISTRIBUTION NOT FOR SALE OR DISTRIBUTION © Jones & Bartlett Learning, LLC © Jones & Bartlett Learning, LLC NOT FOR SALE OR DISTRIBUTION NOT FOR SALE OR DISTRIBUTION © Jones & Bartlett Learning, LLC © Jones & Bartlett Learning, LLC NOT FOR SALE OR DISTRIBUTION NOT FOR SALE OR DISTRIBUTION © Jones & Bartlett Learning, LLC © Jones & Bartlett Learning, LLC NOT FOR SALE OR DISTRIBUTION NOT FOR SALE OR DISTRIBUTION © Jones & Bartlett Learning, LLC © Jones & Bartlett Learning, LLC NOT FOR SALE OR DISTRIBUTION NOT FOR SALE OR DISTRIBUTION © Jones & Bartlett Learning, LLC © Jones & Bartlett Learning, LLC NOT FOR SALE OR DISTRIBUTION NOT FOR SALE OR DISTRIBUTION © Jones & Bartlett Learning, LLC © Jones & Bartlett Learning, LLC NOT FOR SALE OR DISTRIBUTION NOT FOR SALE OR DISTRIBUTION © Jones & Bartlett Learning, LLC © Jones & Bartlett Learning, LLC NOT FOR SALE OR DISTRIBUTION NOT FOR SALE OR DISTRIBUTION © Jones & Bartlett Learning, LLC. NOT FOR SALE OR DISTRIBUTION. 2ND PAGES 9781284032093_CH04_0060.indd 60 8/28/13 12:08 PM CHAPTER 4: Classification of Mammals 61 © Jones Despite& Bartlett their Learning,remarkable success, LLC mammals are much less© Jones stress & onBartlett the taxonomic Learning, aspect LLCof mammalogy, but rather as diverse than are most invertebrate groups. This is probably an attempt to provide students with sufficient information NOT FOR SALE OR DISTRIBUTION NOT FORattributable SALE OR to theirDISTRIBUTION far greater individual size, to the high on the various kinds of mammals to make the subsequent energy requirements of endothermy, and thus to the inabil- discussions of mammalian biology meaningful. -

The Largest Fossil Rodent Andre´S Rinderknecht1 and R
Proc. R. Soc. B doi:10.1098/rspb.2007.1645 Published online The largest fossil rodent Andre´s Rinderknecht1 and R. Ernesto Blanco2,* 1Museo Nacional de Historia Natural y Antropologı´a, Montevideo 11300, Uruguay 2Facultad de Ingenierı´a, Instituto de Fı´sica, Julio Herrera y Reissig 565, Montevideo 11300, Uruguay The discovery of an exceptionally well-preserved skull permits the description of the new South American fossil species of the rodent, Josephoartigasia monesi sp. nov. (family: Dinomyidae; Rodentia: Hystricognathi: Caviomorpha). This species with estimated body mass of nearly 1000 kg is the largest yet recorded. The skull sheds new light on the anatomy of the extinct giant rodents of the Dinomyidae, which are known mostly from isolated teeth and incomplete mandible remains. The fossil derives from San Jose´ Formation, Uruguay, usually assigned to the Pliocene–Pleistocene (4–2 Myr ago), and the proposed palaeoenviron- ment where this rodent lived was characterized as an estuarine or deltaic system with forest communities. Keywords: giant rodents; Dinomyidae; megamammals 1. INTRODUCTION 3. HOLOTYPE The order Rodentia is the most abundant group of living MNHN 921 (figures 1 and 2; Museo Nacional de Historia mammals with nearly 40% of the total number of Natural y Antropologı´a, Montevideo, Uruguay): almost mammalian species recorded (McKenna & Bell 1997; complete skull without left zygomatic arch, right incisor, Wilson & Reeder 2005). In general, rodents have left M2 and right P4-M1. body masses smaller than 1 kg with few exceptions. The largest living rodent, the carpincho or capybara 4. AGE AND LOCALITY (Hydrochoerus hydrochaeris), which lives in the Neotro- Uruguay, Departament of San Jose´, coast of Rı´odeLa pical region of South America, has a body mass of Plata, ‘Kiyu´’ beach (348440 S–568500 W). -

Fossil Calibration Schemes the Soft Explosive Model Of
Supplemental File 2: Fossil calibration schemes The soft explosive model of placental mammal evolution Matthew J. Phillips*,1 and Carmelo Fruciano1 1School of Earth, Environmental and Biological Sciences, Queensland University of Technology, Brisbane, Australia *Corresponding author: E-mail: [email protected] Contents Fossil Calibration schemes ......................................................................................................... 1 Table S2 .................................................................................................................................. 1 New calibrations ..................................................................................................................... 3 Figure S2 ................................................................................................................................ 7 References ............................................................................................................................ 10 Calibration schemes Table S2. Soft-bound calibrations employed for the MCMCtree analyses of the 122-taxon, 128- taxon, and 57-taxon empirical datasets. Calibrations among placental mammals are largely based on dos Reis et al. [1], hence the designations dR32 and dR40 (numbers indicating the number of calibrations). Several new calibrations, including some inspired by Springer et al. [2], are described below the table. Also note that the 122-taxon dR40, 128-taxon and 57-taxon analyses employ bounds as listed below, whereas the dR32 analyses, -

Dental Homologies and Evolutionary Transformations In
Dental homologies and evolutionary transformations in Caviomorpha (Hystricognathi, Rodentia): new data from the Paleogene of Peruvian Amazonia Myriam Boivin, Laurent Marivaux To cite this version: Myriam Boivin, Laurent Marivaux. Dental homologies and evolutionary transformations in Caviomor- pha (Hystricognathi, Rodentia): new data from the Paleogene of Peruvian Amazonia. Historical Biology, Taylor & Francis, 2020, 32 (4), pp.528-554. 10.1080/08912963.2018.1506778. hal-01870927 HAL Id: hal-01870927 https://hal.umontpellier.fr/hal-01870927 Submitted on 17 Jun 2020 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. Page 1 of 118 Historical Biology 1 2 3 Dental homologies and evolutionary transformations in Caviomorpha (Hystricognathi, 4 5 Rodentia): new data from the Paleogene of Peruvian Amazonia 6 7 8 9 10 a* a 11 Myriam Boivin and Laurent Marivaux 12 13 14 15 a Laboratoire de Paléontologie, Institut des Sciences de l’Évolution de Montpellier (ISE-M), c.c. 16 For Peer Review Only 17 18 064, Université de Montpellier, CNRS, IRD, EPHE, place Eugène Bataillon, F-34095 19 20 Montpellier Cedex 05, France. 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 *Corresponding author. -

Investigating Evolutionary Processes Using Ancient and Historical DNA of Rodent Species
Investigating evolutionary processes using ancient and historical DNA of rodent species Thesis submitted for the degree of Doctor of Philosophy (PhD) University of London Royal Holloway University of London Egham, Surrey TW20 OEX Selina Brace November 2010 1 Declaration I, Selina Brace, declare that this thesis and the work presented in it is entirely my own. Where I have consulted the work of others, it is always clearly stated. Selina Brace Ian Barnes 2 “Why should we look to the past? ……Because there is nowhere else to look.” James Burke 3 Abstract The Late Quaternary has been a period of significant change for terrestrial mammals, including episodes of extinction, population sub-division and colonisation. Studying this period provides a means to improve understanding of evolutionary mechanisms, and to determine processes that have led to current distributions. For large mammals, recent work has demonstrated the utility of ancient DNA in understanding demographic change and phylogenetic relationships, largely through well-preserved specimens from permafrost and deep cave deposits. In contrast, much less ancient DNA work has been conducted on small mammals. This project focuses on the development of ancient mitochondrial DNA datasets to explore the utility of rodent ancient DNA analysis. Two studies in Europe investigate population change over millennial timescales. Arctic collared lemming (Dicrostonyx torquatus) specimens are chronologically sampled from a single cave locality, Trou Al’Wesse (Belgian Ardennes). Two end Pleistocene population extinction-recolonisation events are identified and correspond temporally with - localised disappearance of the woolly mammoth (Mammuthus primigenius). A second study examines postglacial histories of European water voles (Arvicola), revealing two temporally distinct colonisation events in the UK. -

Rodentia: Erethizontidae) Therya, Vol
Therya E-ISSN: 2007-3364 [email protected] Asociación Mexicana de Mastozoología México LEON-ALVARADO, OMAR DANIEL; RAMÍREZ-CHAVES, HÉCTOR E. Morphological description of the glans penis and baculum of Coendou quichua (Rodentia: Erethizontidae) Therya, vol. 8, núm. 3, 2017, pp. 263-267 Asociación Mexicana de Mastozoología Baja California Sur, México Available in: http://www.redalyc.org/articulo.oa?id=402352772011 How to cite Complete issue Scientific Information System More information about this article Network of Scientific Journals from Latin America, the Caribbean, Spain and Portugal Journal's homepage in redalyc.org Non-profit academic project, developed under the open access initiative THERYA, 2017, Vol. 8 (3): 263-268 DOI: 10.12933/therya- 17-495 ISSN 2007-3364 Morphological description of the glans penis and baculum of Coendou quichua (Rodentia: Erethizontidae) OMAR D ANIEL LEON-ALVARA DO 1* AN D H ÉCT OR E. RAMÍREZ -CHAVES 2 1 Laboratorio de Sistemática y Biogeografía, Escuela de Biología, Facultad de Ciencias, Universidad Industrial de Santander. Carrera 27 9, A. A. 678, Bucaramanga, Colombia. E-mail: [email protected] (ODLA) 2 Departamento de Ciencias Biológicas, Facultad de Ciencias Exactas y Naturales, Universidad de Caldas. Calle 65 26-10, A. A. 275, Manizales, Colombia. E-mail: [email protected] * Corresponding author External morphology of the glans and baculum are important characters for specic delimitation, especially for rodents (Simson et al. 1995). However, for Erethizontidae there are few descriptive works; in fact, for Neotropical porcupines of the genus Coendou there is just one brief contribution for an indeterminate species by Pocock in 1922. -

Animal Health Requirements for Importation of Rodents, Hedgehogs, Gymnures and Tenrecs Into Denmark
INTERNATIONAL TRADE DIVISION ANIMAL HEALTH REQUIREMENTS FOR IMPORTATION OF RODENTS, HEDGEHOGS, GYMNURES AND TENRECS INTO DENMARK. La 23,0-2111 These animal health requirements concern veterinary import requirements and certification re- quirements alone and shall apply without prejudice to other Danish and EU legislation. Rodents, hedgehogs, gymnures and tenrecs meaning animals of the Genera/Species listed below: Order Family Rodentia Sciuridae (Squirrels) (except Petaurista spp., Biswamoyopterus spp., Aeromys spp., Eupetaurus spp., Pteromys spp., Glaucomys spp., Eoglaucomys spp., Hylopetes spp., Petinomys spp., Aeretes spp., Trogopterus spp., Belomys, Pteromyscus spp., Petaurillus spp., Iomys spp.), Gliridae (Dormous’), Heteromyidae (Kangaroo rats, kangaroo mice and rock pocket mice), Geomyidae (Gophers), Spalaci- dae (Blind mole rats, bamboo rats, root rats, and zokors), Calomyscidae (Mouse-like hamsters), Ne- somyidae (Malagasy rats and mice, climbing mice, African rock mice, swamp mice, pouched rats, and the white-tailed rat), Cricetidae (Hamsters, voles, lemmings, and New World rats and mice), Muridae (mice and rats and gerbils), Dipodidae (jerboas, jumping mice, and birch mice), Pedetidae (Spring- hare), Ctenodactylidae (Gundis), Diatomyidae (Laotian rock rat), Petromuridae (Dassie Rat), Thryon- omyidae (Cane rats), Bathyergidae (Blesmols), Dasyproctidae (Agoutis and acouchis), Agoutidae (Pacas), Dinomyidae (Pacarana), Caviidae (Domestic guinea pig, wild cavies, mara and capybara), Octodontidae (Rock rats, degus, coruros, and viscacha rats), Ctenomyidae (Tuco-tucos), Echimyidae (Spiny rats), Myocastoridae (Coypu ), Capromyidae (Hutias), Chinchillidae (Chinchillas and visca- chas), Abrocomidae (Chinchilla rats). Erinaceomorpha Erinaceidae (Hedgehogs and gymnures) Afrosoricida Tenrecidae (Tenrecs) The importation of rodents, hedgehogs, gymnures and tenrecs to Denmark (excluding import to ap- proved bodies, institutes and centres as defined in Art. 2, 1, (c) of Directive 92/65/EEC) must comply with the requirements of Danish order no. -

How Many Species of Mammals Are There?
Journal of Mammalogy, 99(1):1–14, 2018 DOI:10.1093/jmammal/gyx147 INVITED PAPER How many species of mammals are there? CONNOR J. BURGIN,1 JOCELYN P. COLELLA,1 PHILIP L. KAHN, AND NATHAN S. UPHAM* Department of Biological Sciences, Boise State University, 1910 University Drive, Boise, ID 83725, USA (CJB) Department of Biology and Museum of Southwestern Biology, University of New Mexico, MSC03-2020, Albuquerque, NM 87131, USA (JPC) Museum of Vertebrate Zoology, University of California, Berkeley, CA 94720, USA (PLK) Department of Ecology and Evolutionary Biology, Yale University, New Haven, CT 06511, USA (NSU) Integrative Research Center, Field Museum of Natural History, Chicago, IL 60605, USA (NSU) 1Co-first authors. * Correspondent: [email protected] Accurate taxonomy is central to the study of biological diversity, as it provides the needed evolutionary framework for taxon sampling and interpreting results. While the number of recognized species in the class Mammalia has increased through time, tabulation of those increases has relied on the sporadic release of revisionary compendia like the Mammal Species of the World (MSW) series. Here, we present the Mammal Diversity Database (MDD), a digital, publically accessible, and updateable list of all mammalian species, now available online: https://mammaldiversity.org. The MDD will continue to be updated as manuscripts describing new species and higher taxonomic changes are released. Starting from the baseline of the 3rd edition of MSW (MSW3), we performed a review of taxonomic changes published since 2004 and digitally linked species names to their original descriptions and subsequent revisionary articles in an interactive, hierarchical database. We found 6,495 species of currently recognized mammals (96 recently extinct, 6,399 extant), compared to 5,416 in MSW3 (75 extinct, 5,341 extant)—an increase of 1,079 species in about 13 years, including 11 species newly described as having gone extinct in the last 500 years. -

In the Pleistocene of South America: Biogeographic and Paleoenvironmental Implications
Journal of South American Earth Sciences 82 (2018) 76e90 Contents lists available at ScienceDirect Journal of South American Earth Sciences journal homepage: www.elsevier.com/locate/jsames The southernmost record of a large erethizontid rodent (Hystricomorpha: Erethizontoidea) in the Pleistocene of South America: Biogeographic and paleoenvironmental implications * Raúl I. Vezzosi a, , Leonardo Kerber b a Laboratorio de Paleontología de Vertebrados, Centro de Investigaciones Científicas y Transferencia de Tecnología a la Produccion, Consejo Nacional de Investigaciones Científicas y Tecnicas, Materi y Espana,~ E3105BWA, Diamante, Argentina b CAPPA - Centro de Apoio a Pesquisa Paleontologica da Quarta Colonia,^ Universidade Federal de Santa Maria, Sao~ Joao~ do Pol^esine, Rua Maximiliano Vizzotto, 598, CEP 97230-000, Brazil article info abstract Article history: The South American porcupines (Erethizontidae) are included in two genera: Chaetomys and Coendou. Received 19 October 2017 The latter is a very speciose taxon, with about 13 living species. During at least the late Plioceneeearly Received in revised form Pleistocene, erethizontids immigrated to Central and North America during the Great American Biotic 23 December 2017 Interchange. Although some Pleistocene fossils have been reported, the Quaternary history of this clade Accepted 24 December 2017 is still understudied. The only known extinct species is Coendou magnus. In this work, a fossil of a Available online 30 December 2017 porcupine is reported from an Upper Pleistocene fluvial sedimentary sequence cropping out in the Northern Pampa geomorphological region, Santa Fe Province, Argentina. Despite this group having Keywords: Fossil record different living forms widely distributed in South American Neotropical woodland habitats, the Pleis- Quaternary tocene occurrences of Erethizontidae are scarce and limited to Upper Pleistocene deposits from Bolivia, Regional extinction Brazil, and Uruguay. -
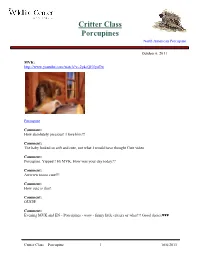
Critter Class Porcupines
Critter Class Porcupines North American Porcupine October 6, 2011 MVK: http://www.youtube.com/watch?v=2y4cQEEyuTw Porcupine Comment: How absolutely precious! I love him!!! Comment: The baby looked so soft and cute, not what I would have thought Cute video Comment: Porcupine. Yippee!! Hi MVK. How was your day today?? Comment: Awwww toooo cute!!! Comment: How cute is that! Comment: OUCH! Comment: Evening MVK and EN - Porcupines - wow - funny little critters or what!!! Good choice♥♥♥ Critter Class – Porcupine 1 10/6/2011 Comment: Oh my goodness -----how cute is that baby porcupine!! I really don't know much about them. He was using his paws so well. Do they have thumbs like raccoons? Comment: Oh my word, isn't that baby cute! And such a dainty eater! Comment: Is that a porcupine? Pulled many of quills from my dogs. Comment: Oh, one of my favorite animals. I love our type, and those snazzy African ones whose head look like they are wearing a fancy Parisian hat. I just love porcupines! MVK: Porcupines have soft hair, but on their back, sides, and tail it is usually mixed with sharp quills. These quills typically lie flat until a porcupine is threatened, then leap to attention as a persuasive deterrent. Porcupines cannot shoot them at predators as once thought, but the quills do detach easily when touched. from National Geographic MVK: http://www.youtube.com/watch?v=wYC0IYuOYLw&NR=1 MVK: http://www.youtube.com/watch?v=mqc5jJQwVu8&feature=relmfu MVK: This little porcupine's mother was hit and killed by a car. -

Early Oligocene Chinchilloid Caviomorphs from Puerto Rico and the Initial Rodent Colonization of the West Indies
Early Oligocene chinchilloid caviomorphs from Puerto Rico and the initial rodent colonization of the West Indies Laurent Marivaux, Jorge Vélez-Juarbe, Gilles Merzeraud, Francois Pujos, Lázaro Viñola López, Myriam Boivin, Hernán Santos-Mercado, Eduardo Cruz, Alexandra Grajales, James Padilla, et al. To cite this version: Laurent Marivaux, Jorge Vélez-Juarbe, Gilles Merzeraud, Francois Pujos, Lázaro Viñola López, et al.. Early Oligocene chinchilloid caviomorphs from Puerto Rico and the initial rodent colonization of the West Indies. Proceedings of the Royal Society B: Biological Sciences, Royal Society, The, 2020, 287, pp.20192806. 10.1098/rspb.2019.2806. hal-02477415 HAL Id: hal-02477415 https://hal.umontpellier.fr/hal-02477415 Submitted on 17 Jun 2020 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. Page 3 of 24 Submitted to Proceedings of the Royal Society B: For Review Only Early Oligocene chinchilloid caviomorphs from Puerto Rico and the initial rodent colonisation of the West Indies Laurent Marivaux1, Jorge Vélez-Juarbe2, Gilles Merzeraud3, François Pujos4, Lázaro W. Viñola López5, Myriam Boivin6, Hernán Santos-Mercado7, Eduardo J. Cruz7, Alexandra Grajales7, James Padilla7, Kevin I. Vélez-Rosado8, Mélody Philippon9, Jean-Len Léticée9, Philippe Münch3 and Pierre-Olivier Antoine1 1Laboratoire de Paléontologie, Institut des Sciences de l’Évolution de Montpellier (ISE-M, UMR 5554, CNRS/UM/IRD/EPHE), c.c.