Download This Article in PDF Format
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Raby Redcurrant Crumble Tray Bake
Raby Redcurrant Crumble tray bake Preparation total time 1 hour Makes 12 portion Nutrition: 100g per serving Kcal Fat Saturates Carbs Sugars Fibre Protein Salt 334 9.41g 5.88g 60.68g 34.92g 1.17g 3.07g 0.40g A layered traybake inspired by German Coffee shops. Ingredients For the berry filling • 200gr Raby red currants • 50g Caster Sugar • 2 tbsp Corn flour For the Cake • 220g Plain Flour • 4g Baking Powder • 2g Cinnamon • 50g Ground Almonds • 200g Caster Sugar • 80g Butter, Melted • 2 Free- range eggs • 40g Yoghurt- thinned with 2tbs milk Method 1. Preheat the oven to 180C/350F/Gas 4. Grease and line a rectangular baking tin (approximately 26cm x 20cm/10½in x 8in). 2. In a saucepan combine the fruit with 50ml/2fl oz water. Bring the fruit just to the boil, reduce the heat and simmer for 2-3 minutes. 3. In a mixing bowl combine the caster sugar with the cornflour. Stir into the fruit and continue to cook for 2-3 minutes, stirring frequently, until the mixture is thick and jammy. Set aside to cool. 4. For the crumble topping, add the flour to a mixing bowl and rub in the butter until the mixture resembles fine breadcrumbs and no large lumps of butter are left. Stir in the sugar and set aside. 5. For the cake, in a large mixing bowl, sift together the flour, baking powder and cinnamon. Stir in the ground almonds until thoroughly combined. 6. In another bowl beat together the sugar with the melted butter, eggs, yoghurt and milk mixture until thoroughly mixed. -

Provision of Information on the Amount of Sugar in Processed Foods
DOI: 10.1590/1413-81232021263.07872019 1153 Provision of information on the amount of sugar FREE THEMES in processed foods Camila Cremonezi Japur (https://orcid.org/0000-0003-0513-1758) 1,4 Dyessa Cardoso Bernardes Assunção (https://orcid.org/0000-0003-4711-7508) 2 Raíssa Aparecida Borges Batista (https://orcid.org/0000-0002-9928-5872) 2 Fernanda Rodrigues de Oliveira Penaforte (https://orcid.org/0000-0001-8483-1562) 3,4 Abstract The objective of this study was to assess the provision of information on the amount of sugar and identify the position of sugar in the list of ingredients of processed foods. A cross-sectional study was conducted to analyze all processed tra- ditional and diet/light/zero food products sold in a hypermarket containing the word sugar or sucrose in the list of ingredients. The food labels were read and the position of sugar on the list of ingredients and presence, or absence, of information on the amount of sugar in the nutrition facts table were 1 Divisão de Nutrição recorded. Information on the amount of sugar was e Metabolismo, Departamento de Ciências also requested from the manufacturers by e-mail da Saúde, Faculdade de or telephone. A total of 2,200 food products were Medicina de Ribeirão Preto, assessed, 2,164 (98.4%) of which were tradition- Universidade de São Paulo (USP). Av. Bandeirantes al foods and 36 (1.6%) diet/light/zero foods. The 3900, Monte Alegre. 14049- amount of sugar was declared in only 14.4% and 900 Ribeirão Preto SP 13.9% of these products, respectively (p=0.84). -
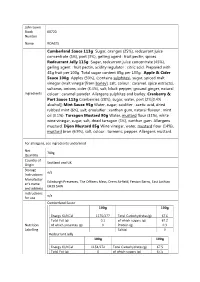
Cumberland Sauce 113G Sugar, Oranges (25%), Redcurrant Juice Concentrate (5%), Port (3%), Gelling Agent : Fruit Pectin, Spices
John Lewis Stock 60720 Number Name ROASTS Cumberland Sauce 113g Sugar, oranges (25%), redcurrant juice concentrate (5%), port (3%), gelling agent : fruit pectin, spices. Redcurrant Jelly 113g Sugar, redcurrant juice concentrate (45%), gelling agent : fruit pectin, acidity regulator : citric acid. Prepared with 45g fruit per 100g. Total sugar content 65g per 100g. Apple & Cider Sauce 100g Apples (50%), (contains sulphites), sugar, spiced malt vinegar (malt vinegar[from barley], salt, colour : caramel, spice extracts), sultanas, onions, cider (3.4%), salt, black pepper, ground ginger, natural Ingredients colour : caramel powder. Allergens sulphites and barley. Cranberry & Port Sauce 113g Cranberries (38%), sugar, water, port (2%[0.4% alcohol]) Mint Sauce 95g Water, sugar, acidifier : acetic acid, dried rubbed mint (6%), salt, emulsifier : xanthan gum, natural flavour : mint oil (0.1%). Tarragon Mustard 90g Water, mustard flour (31%), white wine vinegar, sugar, salt, dried tarragon (3%), xanthan gum. Allergens mustard. Dijon Mustard 85g Wine vinegar, water, mustard flour (14%), mustard bran (6.9%), salt, colour : turmeric, pepper. Allergens mustard. For allergens, see ingredients underlined Net 709g Quantity Country of Scotland and UK Origin Storage n/a Instructions Manufactur Edinburgh Preserves, The Officers Mess, Drem Airfield, Fenton Barns, East Lothian er’s name EH39 5AW and address Instructions n/a for use Cumberland Sauce 100g 100g Energy KJ/KCal 1175/277 Total Carbohydrates(g) 67.6 Total Fat (g) 0.1 of which sugars (g) 67.2 Nutrition -

WSET Level 2 Systematic Approach to Tasting Wine®
Level 2 SAT (A5) 2012 (v2)_Layout 1 30/03/2012 15:47 Page 1 WSET Level 2 Systematic Approach to Tasting Wine ® APPEARANCE Clarity clear – hazy Intensity pale – medium – deep Colour white lemon – gold – amber rosé pink – salmon – orange red purple – ruby – garnet – tawny NOSE Condition clean – unclean Intensity light – medium – pronounced Aroma characteristics e.g. fruits, flowers, spices, vegetables, oak aromas, other PALATE Sweetness dry – off-dry – medium – sweet Acidity low – medium – high Tannin low – medium – high Body light – medium – full Flavour characteristics e.g. fruits, flowers, spices, vegetables, oak flavours, other Finish short – medium – long CONCLUSIONS Quality faulty – poor – acceptable – good – very good – outstanding Copyright Wine & Spirit Education Trust 2012. The WSET Level 2 Systematic Approach to Tasting Wine ® may only be reproduced with the written permission of the WSET subject to their terms and conditions. For more information contact [email protected] 2W-Version 1.0 Level 2 SAT (A5) 2012 (v2)_Layout 1 30/03/2012 15:47 Page 2 WSET Level 2 Wine-Lexicon: supporting the WSET Level 2 Systematic Approach to Tasting Wine ® AROMA AND FLAVOUR CHARACTERISTICS FLORAL / FRUIT Are the flavours simple/generic or specific? Fresh or cooked? Ripe or unripe? Floral blossom, rose, violet Green Fruit green apple, red apple, gooseberry, pear, grape Citrus Fruit grapefruit, lemon, lime (juice or zest?) Stone Fruit peach, apricot, nectarine Tropical Fruit banana, lychee, mango, melon, passion fruit, pineapple Red Fruit redcurrant, -

Well-Known Plants in Each Angiosperm Order
Well-known plants in each angiosperm order This list is generally from least evolved (most ancient) to most evolved (most modern). (I’m not sure if this applies for Eudicots; I’m listing them in the same order as APG II.) The first few plants are mostly primitive pond and aquarium plants. Next is Illicium (anise tree) from Austrobaileyales, then the magnoliids (Canellales thru Piperales), then monocots (Acorales through Zingiberales), and finally eudicots (Buxales through Dipsacales). The plants before the eudicots in this list are considered basal angiosperms. This list focuses only on angiosperms and does not look at earlier plants such as mosses, ferns, and conifers. Basal angiosperms – mostly aquatic plants Unplaced in order, placed in Amborellaceae family • Amborella trichopoda – one of the most ancient flowering plants Unplaced in order, placed in Nymphaeaceae family • Water lily • Cabomba (fanwort) • Brasenia (watershield) Ceratophyllales • Hornwort Austrobaileyales • Illicium (anise tree, star anise) Basal angiosperms - magnoliids Canellales • Drimys (winter's bark) • Tasmanian pepper Laurales • Bay laurel • Cinnamon • Avocado • Sassafras • Camphor tree • Calycanthus (sweetshrub, spicebush) • Lindera (spicebush, Benjamin bush) Magnoliales • Custard-apple • Pawpaw • guanábana (soursop) • Sugar-apple or sweetsop • Cherimoya • Magnolia • Tuliptree • Michelia • Nutmeg • Clove Piperales • Black pepper • Kava • Lizard’s tail • Aristolochia (birthwort, pipevine, Dutchman's pipe) • Asarum (wild ginger) Basal angiosperms - monocots Acorales -

Ribes Growers' Guide
RIBES GROWERS’ GUIDE (2013) Table Of Contents Introduction 2 Black currants Varieties 3 Planting 6 Preparation 6 Site Requirements 6 Spacing 6 Pruning 7 Hand Pruning 7 Mechanical Pruning 8 Fertilizer 8 Weed Control 9 Pests 9 Aphids 10 Currant Borer 10 Currant Fruit Fly 10 Mites 10 Anthracnose, Leaf Spot 10 Powdery Mildew 11 White Pine Blister Rust 11 Viruses 11 Water Management 12 Red Currants Varieties 13 Red Varieties 13 White Varieties 14 Planting 14 Spacing 14 Pruning 15 Bush Form 15 Cordons 15 Pests 16 Aphids 16 Sawfly 16 Gooseberries Varieties 16 Planting 16 Pruning 16 1 Introduction This is the sixteenth annual McGinnis Berry Crops guide to production of Ribes plant varieties. The purpose of the guide is to provide the necessary information about variety selection, planting requirements, and plant care that growers will need to make informed decisions. This guide will look at blackcurrants, red currants (including white and pink, which are of the same species as reds), and gooseberries. While not all varieties are included in this guide, we cover those that seem to have the most potential for the North American market. Common varieties such as Pixwell, Oregon Champion, Colossal (gooseberries), Consort, Ben Nevis, Crandall, Ben Lomond, Ben Alder (black currants) have been surpassed by the introduction of varieties that combine higher yields, better disease resistance and superior fruit qualities. In future editions of this guide we will discuss new and relevant varieties as they are made available, as well as updated information on yields, pests, and information from the plant trials that will be taking place across North America. -

Download Authenticated
Ohio Administrative Code Rule 1501:18-1-03 Endangered and threatened species. Effective: January 30, 2021 (A) The following species of plants are designated as endangered in Ohio. (1) Acer pensylvanicum L., Striped maple. (2) Aconitum noveboracense A. Gray, Northern monkshood. (3) Aconitum uncinatum L., Southern monkshood. (4) Adlumia fungosa (Ait.) Greene, Allegheny-vine. (5) Agalinis auriculata (Michx.) Blake, Ear-leaved-foxglove. (6) Agalinis purpurea (L.) Pennell var. parviflora (Benth.) Boivin, Small purple-foxglove. (7) Agalinis skinneriana (Wood) Britt., Skinner's-foxglove. (8) Ageratina aromatica (L.) Spach, Small white snakeroot. (9) Agrostis elliottiana Schultes, Elliott's bent grass. (10) Amelanchier humilis Wiegand, Low serviceberry. (11) Amelanchier interior E.L. Nielsen, Inland serviceberry. (12) Amphidium mougeotii (Bruch, Schimper and W. Gmbel) Schimper, Mougeot's ice moss. (13) Andropogon glomeratus (Walter) Britton, Bushy broom-sedge. Page 1 (14) Androsace occidentalis Pursh, Western rock-jasmine. (15) Anomobryum filiforme (Dicks.) Solms, Common silver moss. (16) Anomodon viticulosus (Hedw.) Hook. and Taylor, Long tail moss. (17) Arabidopsis lyrata (L.) OKane and Al-Shehbaz, Lyre-leaved rock cress. (18) Arabis patens Sullivant, Spreading rock cress. (19) Arctostaphylos uva-ursi (L.) Spreng., Bearberry. (20) Aralia hispida Vent., Bristly sarsaparilla. (21) Arethusa bulbosa L., Dragon's-mouth. (22) Aristida basiramea Engelm. ex Vasey, Forked three-awn grass. (23) Aristida necopina Shinners, False arrow-feather. (24) Aronia arbutifolia (L.) Pers., Red chokeberry. (25) Asplenium bradleyi D.C. Eaton, Bradley's spleenwort. (26) Asplenium resiliens Kunze, Black-stemmed spleenwort. (27) Astragalus neglectus (T. and G.) Sheld., Cooper's milk-vetch. (28) Baptisia australis (L.) R. Br., Blue false indigo. (29) Barbula indica (Hooker) Sprengel in E.G. -

Acute Supplementation with Blackcurrant Extracts Modulates Cognitive Functioning and Inhibits Monoamine Oxidase-B in Healthy Young Adults
Journal of Functional Foods 17 (2015) 524–539 Available online at www.sciencedirect.com ScienceDirect journal homepage: www.elsevier.com/locate/jff Acute supplementation with blackcurrant extracts modulates cognitive functioning and inhibits monoamine oxidase-B in healthy young adults Anthony W. Watson a,b,*, Crystal F. Haskell-Ramsay a, David O. Kennedy a, Janine M. Cooney b, Tania Trower b, Arjan Scheepens b a Brain, Performance and Nutrition Research Centre, Northumbria University, Newcastle Upon-Tyne NE1 8ST, UK b The New Zealand Institute for Plant & Food Research Ltd, Auckland 1025, New Zealand ARTICLE INFO ABSTRACT Article history: The consumption of berry fruits engenders a number of benefits in animal models, includ- Received 27 February 2015 ing improvements in cognitive performance, slowing of cognitive decline during natural ageing, Received in revised form 2 June and neuroprotection. These findings, along with limited human epidemiological evidence, 2015 suggest a potential role for the consumption of berry fruit polyphenols in improving human Accepted 4 June 2015 cognitive performance. The current study assessed the effects of two blackcurrant extracts Available online on cognitive outcomes, mood, autonomic measures, peripheral and central monoamine tone, and anthocyanin bioavailability to plasma. A randomised, double-blind, placebo-controlled, Keywords: crossover study was conducted using 36 healthy young participants (18–35 years). Findings Blackcurrants from the intervention illustrate a cognitive benefit of acute blackcurrant -

Fats and Fatty Acid in Human Nutrition
ISSN 0254-4725 91 FAO Fats and fatty acids FOOD AND NUTRITION PAPER in human nutrition Report of an expert consultation 91 Fats and fatty acids in human nutrition − Report of an expert consultation Knowledge of the role of fatty acids in determining health and nutritional well-being has expanded dramatically in the past 15 years. In November 2008, an international consultation of experts was convened to consider recent scientific developments, particularly with respect to the role of fatty acids in neonatal and infant growth and development, health maintenance, the prevention of cardiovascular disease, diabetes, cancers and age-related functional decline. This report will be a useful reference for nutrition scientists, medical researchers, designers of public health interventions and food producers. ISBN 978-92-5-106733-8 ISSN 0254-4725 9 7 8 9 2 5 1 0 6 7 3 3 8 Food and Agriculture I1953E/1/11.10 Organization of FAO the United Nations FAO Fats and fatty acids FOOD AND NUTRITION in human nutrition PAPER Report of an expert consultation 91 10 − 14 November 2008 Geneva FOOD AND AGRICULTURE ORGANIZATION OF THE UNITED NATIONS Rome, 2010 The designations employed and the presentation of material in this information product do not imply the expression of any opinion whatsoever on the part of the Food and Agriculture Organization of the United Nations (FAO) concerning the legal or development status of any country, territory, city or area or of its authorities, or concerning the delimitation of its frontiers or boundaries. The mention of specific companies or products of manufacturers, whether or not these have been patented, does not imply that these have been endorsed or recommended by FAO in preference to others of a similar nature that are not mentioned. -

Blackcurrant Variety Specific Growth and Yield Formation As a Response
agronomy Article Blackcurrant Variety Specific Growth and Yield Formation as a Response to Foliar Fertilizers 1 2 1 3 4, Sorin Vâtcă , Roxana Vidican ,S, tefania Gâdea , Melinda Horvat , Anamaria Vâtcă *, 1, 2 Valentina Ancut, a Stoian * and Vlad Stoian 1 Department of Plant Physiology, Faculty of Agriculture, University of Agricultural Sciences and Veterinary Medicine Cluj-Napoca, Calea Mănă¸stur3–5, 400372 Cluj-Napoca, Romania; [email protected] (S.V.); [email protected] (S, .G.) 2 Department of Microbiology, Faculty of Agriculture, University of Agricultural Sciences and Veterinary Medicine Cluj-Napoca, Calea Mănă¸stur3–5, 400372 Cluj-Napoca, Romania; [email protected] (R.V.); [email protected] (V.S.) 3 Department of Infectious Diseases, Faculty of Medicine, University of Medicine and Pharmacy Iuliu Ha¸tieganuCluj-Napoca, Babe¸sStreet 8, 400012 Cluj-Napoca, Romania; [email protected] 4 Department of Management and Economics, Faculty of Animal Science and Biotechnologies, University of Agricultural Sciences and Veterinary Medicine Cluj-Napoca, Calea Mănă¸stur3–5, 400372 Cluj-Napoca, Romania * Correspondence: [email protected] (A.V.); [email protected] (V.A.S.) Received: 9 November 2020; Accepted: 16 December 2020; Published: 21 December 2020 Abstract: Recently, there has been a growing interest in supplementing plant nutrition with foliar fertilizers. Foliar application of nutrients is considered a promising environmentally friendly strategy for improving the yield and nutrient efficiency of crops. Little work with foliar appliances on perennial fruits bushes, blackcurrant (Ribes nigrum L.), respectively, has been reported. This study was conducted to evaluate the growth and yield response of three blackcurrant varieties to macro and micronutrients’ foliar fertilizer solutions. -

Annotated Checklist of Vascular Flora, Bryce
National Park Service U.S. Department of the Interior Natural Resource Program Center Annotated Checklist of Vascular Flora Bryce Canyon National Park Natural Resource Technical Report NPS/NCPN/NRTR–2009/153 ON THE COVER Matted prickly-phlox (Leptodactylon caespitosum), Bryce Canyon National Park, Utah. Photograph by Walter Fertig. Annotated Checklist of Vascular Flora Bryce Canyon National Park Natural Resource Technical Report NPS/NCPN/NRTR–2009/153 Author Walter Fertig Moenave Botanical Consulting 1117 W. Grand Canyon Dr. Kanab, UT 84741 Sarah Topp Northern Colorado Plateau Network P.O. Box 848 Moab, UT 84532 Editing and Design Alice Wondrak Biel Northern Colorado Plateau Network P.O. Box 848 Moab, UT 84532 January 2009 U.S. Department of the Interior National Park Service Natural Resource Program Center Fort Collins, Colorado The Natural Resource Publication series addresses natural resource topics that are of interest and applicability to a broad readership in the National Park Service and to others in the management of natural resources, including the scientifi c community, the public, and the NPS conservation and environmental constituencies. Manuscripts are peer-reviewed to ensure that the information is scientifi cally credible, technically accurate, appropriately written for the intended audience, and is designed and published in a professional manner. The Natural Resource Technical Report series is used to disseminate the peer-reviewed results of scientifi c studies in the physical, biological, and social sciences for both the advancement of science and the achievement of the National Park Service’s mission. The reports provide contributors with a forum for displaying comprehensive data that are often deleted from journals because of page limitations. -

Floristic Quality Assessment Report
FLORISTIC QUALITY ASSESSMENT IN INDIANA: THE CONCEPT, USE, AND DEVELOPMENT OF COEFFICIENTS OF CONSERVATISM Tulip poplar (Liriodendron tulipifera) the State tree of Indiana June 2004 Final Report for ARN A305-4-53 EPA Wetland Program Development Grant CD975586-01 Prepared by: Paul E. Rothrock, Ph.D. Taylor University Upland, IN 46989-1001 Introduction Since the early nineteenth century the Indiana landscape has undergone a massive transformation (Jackson 1997). In the pre-settlement period, Indiana was an almost unbroken blanket of forests, prairies, and wetlands. Much of the land was cleared, plowed, or drained for lumber, the raising of crops, and a range of urban and industrial activities. Indiana’s native biota is now restricted to relatively small and often isolated tracts across the State. This fragmentation and reduction of the State’s biological diversity has challenged Hoosiers to look carefully at how to monitor further changes within our remnant natural communities and how to effectively conserve and even restore many of these valuable places within our State. To meet this monitoring, conservation, and restoration challenge, one needs to develop a variety of appropriate analytical tools. Ideally these techniques should be simple to learn and apply, give consistent results between different observers, and be repeatable. Floristic Assessment, which includes metrics such as the Floristic Quality Index (FQI) and Mean C values, has gained wide acceptance among environmental scientists and decision-makers, land stewards, and restoration ecologists in Indiana’s neighboring states and regions: Illinois (Taft et al. 1997), Michigan (Herman et al. 1996), Missouri (Ladd 1996), and Wisconsin (Bernthal 2003) as well as northern Ohio (Andreas 1993) and southern Ontario (Oldham et al.