Post-Fire Mortality and Response in a Redwood/ Douglas-Fir Forest
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Local Hazard Mitigation Plan Five Year Update 2017–2022
CITY OF SANTA CRUZ Local Hazard Mitigation Plan Five Year Update 2017–2022 Hazard Mitigation is any action taken to reduce or eliminate the long-term risk to human life and property from hazards. ~ Title 44 Code of Federal Regulations (§206.401) Adopted by the City Council xxxx xx, 2017 Table of Contents APPENDICES .......................................................................................................................................................... II MAPS AND FIGURES ................................................................................................................................................ II TABLES ................................................................................................................................................................ III HOW TO USE THIS PLAN ......................................................................................................................................... IV PART 1 — INTRODUCTION AND ADOPTION .................................................................................................. 1 INTRODUCTION ..................................................................................................................................................... 2 ACKNOWLEDGEMENTS ............................................................................................................................................ 4 SUMMARY ........................................................................................................................................................... -

Santa Cruz County San Mateo County
Santa Cruz County San Mateo County COMMUNITY WILDFIRE PROTECTION PLAN Prepared by: CALFIRE, San Mateo — Santa Cruz Unit The Resource Conservation District for San Mateo County and Santa Cruz County Funding provided by a National Fire Plan grant from the U.S. Fish and Wildlife Service through the California Fire Safe Council. M A Y - 2 0 1 0 Table of Contents Executive Summary.............................................................................................................1 Purpose.................................................................................................................................2 Background & Collaboration...............................................................................................3 The Landscape .....................................................................................................................6 The Wildfire Problem ..........................................................................................................8 Fire History Map................................................................................................................10 Prioritizing Projects Across the Landscape .......................................................................11 Reducing Structural Ignitability.........................................................................................12 x Construction Methods............................................................................................13 x Education ...............................................................................................................15 -

Community Wildfire Protection Plan Prepared By
Santa Cruz County San Mateo County COMMUNITY WILDFIRE PROTECTION PLAN Prepared by: CALFIRE, San Mateo — Santa Cruz Unit The Resource Conservation District for San Mateo County and Santa Cruz County Funding provided by a National Fire Plan grant from the U.S. Fish and Wildlife Service through the California Fire Safe Council. APRIL - 2 0 1 8 Table of Contents Executive Summary ............................................................................................................ 1 Purpose ................................................................................................................................ 3 Background & Collaboration ............................................................................................... 4 The Landscape .................................................................................................................... 7 The Wildfire Problem ........................................................................................................10 Fire History Map ............................................................................................................... 13 Prioritizing Projects Across the Landscape .......................................................................14 Reducing Structural Ignitability .........................................................................................16 • Construction Methods ........................................................................................... 17 • Education ............................................................................................................. -

Fall 2016 1 Chief’S Message
Report to the Community Fall 2016 1 Chief’s Message As Fire Chief of the Albany Fire Department, it is my honor to present the Albany Fire Department’s Report to the Community. In our response to emergencies and our provision of health, safety and emergency services to the community, the department continues to exemplify the excellence and professionalism Albany has come to expect. Although you probably know us as the “fire department,” this by no means reflects the many ways our department serves the community every day. The Albany Fire Department not only responds to fires, but also provides the necessary training to respond to a multitude of emergencies. We also provide emergency medical services including transport, hazardous material response, and technical rescues for vehicle accidents and water rescue. Thanks in great part to the success of our Fire Prevention program, emergency medical calls now account for a majority of our workload. As we move forward into 2017 and beyond, we continue to emphasize training and best practices in everything we do to keep Albany safe. I would like to thank the Mayor, the City Council and the City Manager Mission for their support throughout some tough years, and for their support and vision for AFD’s continued growth and change. The Albany Fire Department enhances the quality of life and the environment by providing fire prevention and suppression, The future of the Albany Fire Department looks tremendous. It is my emergency medical services, public education and emergency honor to be a part of this department and its long, rich history and preparedness. -

Damage and Mortality Assessment of Redwood and Mixed Conifer Forest Types in Santa Cruz County Following Wildfire
Damage and Mortality Assessment of Redwood and Mixed Conifer Forest Types in Santa Cruz County Following Wildfire Steve R. Auten1 and Nadia Hamey1 Abstract On August 12, 2009, the Lockheed Fire ignited the west slope of the Santa Cruz Mountains burning approximately 7,819 acres. A mixture of vegetation types were in the path of the fire, including approximately 2,420 acres of redwood forest and 1,951 acres of mixed conifer forest types representative of the Santa Cruz Mountains. Foresters and land managers were left with tough decisions on how to treat tree damage and mortality compounded by the Pine Mountain Fire which occurred in the same area in 1948. Big Creek Lumber Company (BCL), Cal Poly’s Swanton Pacific Ranch (SPR) and other professionals familiar with this region of redwood teamed up to develop a method for evaluating damage and mortality. Qualitative criteria for evaluating stand damage focused on historic defect, cambial death, root damage, and associated fire intensity. Quantitative damage criteria was used to contrive three mortality assessment tables, broken up by diameter class (1 through 8, 9 through 16, 17+), for all tree species and tested against 83, 1/5th acre fixed plots from SPR’s Continuous Forest Inventory. Since the initial mortality evaluation using the new tables in fall of 2009, each of the 2877 trees have been re-evaluated in spring 2010 and spring 2011. Accuracy against the initial evaluation is 89.3 percent. Key words: damage, hardwood, mortality, redwood Introduction What should be harvested to encourage regeneration of selectively-managed forestland in the Southern Subdistrict of the Coast Forest District following wildfire? What determines tree mortality for the purpose of amending the sustainability analysis (SA) of a Non-industrial Timber Management Plan (NTMP) following wildfire? Charged with managing and maintaining the health and vigor of the forest ecosystem, foresters and land managers need an accurate way of field-evaluating damage and mortality in conifers and hardwoods immediately following wildfire. -

Abstracts and Presenter Biographical Information Oral Presentations
ABSTRACTS AND PRESENTER BIOGRAPHICAL INFORMATION ORAL PRESENTATIONS Abstracts for oral presentations and biographical information for presenters are listed alphabetically below by presenting author’s last name. Abstracts and biographical information appear unmodified, as submitted by the corresponding authors. Day, time, and room number of presentation are also provided. Abatzoglou, John John Abatzoglou, Assistant Professor of Geography, University of Idaho. Research interests span the weather-climate continuum and both basic and applied scientific questions on past, present and future climate dynamics as well as their influence on wildfire, ecology and agriculture and is a key player in the development of integrated climate scenarios for the Pacific Northwest, US. Oral presentation, Wednesday, 2:30 PM, B114 Will climate change increase the occurrence of megafires in the western United States? The largest wildfires in the western United States account for a substantial portion of annual area burned and are associated with numerous direct and indirect geophysical impacts in addition to commandeering suppression resources and national attention. While substantial prior work has been devoted to understand the influence of climate, and weather on annual area burned, there has been limited effort to identify factors that enable and drive the very largest wildfires, or megafires. We hypothesize that antecedent climate and shorter-term biophysically relevant meteorological variables play an essen- tial role in favoring or deterring historical megafire occurrence identified using the Monitoring Trends in Burn Severity Atlas from 1984-2010. Antecedent climatic factors such as drought and winter and spring temperature were found to vary markedly across geographic areas, whereas regional commonality of prolonged extremely low fuel moisture and high fire danger prior to and immediately following megafire discovery. -

Lockheed Fire Post Fire Risk Assessment
Lockheed Fire Post Fire Risk Assessment San Mateo-Santa Cruz Unit California Department of Forestry and Fire Protection September 30, 2009 Aerial photo of Mill Creek drainage August 25, 2009 Table of Contents Page # Executive Summary 1 Introduction and Incident History 1 Watersheds 3 Field Observations 3 Burn Severity Characteristics 8 Fire Behavior 8 Historic Fire Return Intervals 9 Seasonal Context 10 Geology 12 General Observations 13 County Road 13 Downstream Impacts 13 Description of Geologic Units 16 Soils 17 Hydrophobic Soil Conditions 19 Soil Erosion Potential 20 Ecological Impacts 21 Redwood Forest 25 Mixed Conifer 25 Chaparral 26 Coastal Scrub 27 Grassland 27 Specific/Isolated Forest Stands 28 Invasive Species 29 General Recommendations 31 Recommendations to Mitigate Potential Soil Erosion and the Impacts 33 of Storm Water Runoff/Flooding Specific Observations 37 References 51 Post Fire Restoration Do’s and Don’ts 54 Firescaping and Erosion Control with Native Plants 59 Sample No Trespassing Sign 64 Maps Page # Watershed Map 6 Burn Severity Map 7 Fire History Map 11 Geology Map 15 Soil Map 18 Vegetation Type Map 24 Risk Maps Archibald Creek 40 Bettencourt Creek 41 Big Creek 42 Boyer Creek 43 Little Creek 44 Lower Scotts Creek 45 Mill Creek 46 Molino Creek 47 Queseria Creek 48 Upper Scotts Creek 49 Winter Creek 50 Tables and Charts Page # Table 1. Percent of Watersheds Burned 3 Table 2. Ranked Watersheds 5 Table 3. Fire Behavior 9 Table 4. Burn Severity by Vegetation Type 22 Chart 1. Lockheed Fire Severity by Vegetation Type 23 Chart 2. Vegetation Types by Burn Severity 23 Table 5. -
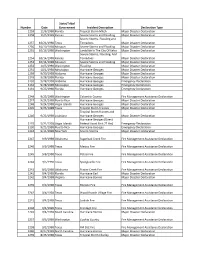
Identification of Disaster Code Declaration
State/Tribal Number Date Government Incident Description Declaration Type 1259 11/6/1998 Florida Tropical Storm Mitch Major Disaster Declaration 1258 11/5/1998 Kansas Severe Storms and Flooding Major Disaster Declaration Severe Storms, Flooding and 1257 10/21/1998 Texas Tornadoes Major Disaster Declaration 1256 10/19/1998 Missouri Severe Storms and Flooding Major Disaster Declaration 1255 10/16/1998 Washington Landslide In The City Of Kelso Major Disaster Declaration Severe Storms, Flooding, And 1254 10/14/1998 Kansas Tornadoes Major Disaster Declaration 1253 10/14/1998 Missouri Severe Storms and Flooding Major Disaster Declaration 1252 10/5/1998 Washington Flooding Major Disaster Declaration 1251 10/1/1998 Mississippi Hurricane Georges Major Disaster Declaration 1250 9/30/1998 Alabama Hurricane Georges Major Disaster Declaration 1249 9/28/1998 Florida Hurricane Georges Major Disaster Declaration 3133 9/28/1998 Alabama Hurricane Georges Emergency Declaration 3132 9/28/1998 Mississippi Hurricane Georges Emergency Declaration 3131 9/25/1998 Florida Hurricane Georges Emergency Declaration 2248 9/25/1998 Washington Columbia County Fire Management Assistance Declaration 1247 9/24/1998 Puerto Rico Hurricane Georges Major Disaster Declaration 1248 9/24/1998 Virgin Islands Hurricane Georges Major Disaster Declaration 1245 9/23/1998 Texas Tropical Storm Frances Major Disaster Declaration Tropical Storm Frances and 1246 9/23/1998 Louisiana Hurricane Georges Major Disaster Declaration Hurricane Georges (Direct 3129 9/21/1998 Virgin Islands Federal -

Local Hazard Mitigation Plan Five Year Update 2018–2023
CITY OF SANTA CRUZ Local Hazard Mitigation Plan Five Year Update 2018–2023 Hazard Mitigation is any action taken to reduce or eliminate the long-term risk to human life and property from hazards. ~ Title 44 Code of Federal Regulations (§206.401) Adopted by the City Council xxxx xx, 201x Table of Contents APPENDICES .......................................................................................................................................................... II MAPS AND FIGURES ................................................................................................................................................ II TABLES ................................................................................................................................................................ III HOW TO USE THIS PLAN ......................................................................................................................................... IV PART 1 — INTRODUCTION AND ADOPTION .................................................................................................... 1 INTRODUCTION...................................................................................................................................................... 2 ACKNOWLEDGEMENTS ............................................................................................................................................ 4 SUMMARY ........................................................................................................................................................... -

015-2018 SHMP FINAL Appendices
APPENDICES – 2018 STATE HAZARD MITIGATION TEAM ROSTER OF AGENCIES AND STAKEHOLDER ORGANIZATIONS AECOM National Governments Alameda County Office of Emergency Services Alpine County Operational Area Inland Region IV Air Resources Control Board (ARB) Amador County Operational Area Inland Region IV American Planning Association California Chapter American Red Cross (Sacramento Chapter) Association of Bay Area Governments Association of Contingency Planners Association of Environmental Professionals Burbank Fire Corps Business and Industry Council for Emergency Planning & Preparedness (BICEPP) Business, Consumer Services, and Housing Agency Business Executives for National Security (BENS) Business Recovery Managers Association Butte County Operational Area Inland Region III California Adaptation Forum California Association of Councils of Governments Cahuilla Band of Indians California Board of Forestry and Fire Protection Calaveras Council of Governments California Coastal Commission California Conservation Corps California Community Colleges California Department of Community Services and Development California Department of Conservation California Department of Corrections and Rehabilitation California Department of Education California Department of Food and Agriculture California Department of Forestry and Fire Protection (CALFIRE) California Department of General Services California Department of Housing and Community Development California Department of Insurance California Department of Public Health California Department of Public -

Community Wildfire Protection Plan
Santa Cruz County San Mateo County COMMUNITY WILDFIRE PROTECTION PLAN Prepared by: CALFIRE, San Mateo — Santa Cruz Unit The Resource Conservation District for San Mateo County and Santa Cruz County Funding provided by a National Fire Plan grant from the U.S. Fish and Wildlife Service through the California Fire Safe Council. M A Y - 2 0 1 0 Table of Contents Executive Summary.............................................................................................................1 Purpose.................................................................................................................................2 Background & Collaboration...............................................................................................3 The Landscape .....................................................................................................................6 The Wildfire Problem ..........................................................................................................8 Fire History Map................................................................................................................10 Prioritizing Projects Across the Landscape .......................................................................11 Reducing Structural Ignitability.........................................................................................12 • Construction Methods............................................................................................13 • Education ...............................................................................................................15 -

SCCO Master Plan
Santa Cruz County Master Fire Plan 2012/2015 Contents EXECUTIVE SUMMARY...........................................................................................................................................2 FINANCIAL ..................................................................................................................................................................6 MANAGEMENT ..........................................................................................................................................................8 VOLUNTEERS..........................................................................................................................................................10 JURISDICTION .........................................................................................................................................................13 FACILITIES................................................................................................................................................................14 VEHICLE REPLACEMENT PROGRAM ...............................................................................................................15 FIRE PREVENTION.................................................................................................................................................17 TRAINING..................................................................................................................................................................19 DISPATCH SERVICES............................................................................................................................................22