Proceedings of the Biological Society of Washington
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Other Contributions
Other Contributions NATURE NOTES Amphibia: Caudata Ambystoma ordinarium. Predation by a Black-necked Gartersnake (Thamnophis cyrtopsis). The Michoacán Stream Salamander (Ambystoma ordinarium) is a facultatively paedomorphic ambystomatid species. Paedomorphic adults and larvae are found in montane streams, while metamorphic adults are terrestrial, remaining near natal streams (Ruiz-Martínez et al., 2014). Streams inhabited by this species are immersed in pine, pine-oak, and fir for- ests in the central part of the Trans-Mexican Volcanic Belt (Luna-Vega et al., 2007). All known localities where A. ordinarium has been recorded are situated between the vicinity of Lake Patzcuaro in the north-central portion of the state of Michoacan and Tianguistenco in the western part of the state of México (Ruiz-Martínez et al., 2014). This species is considered Endangered by the IUCN (IUCN, 2015), is protected by the government of Mexico, under the category Pr (special protection) (AmphibiaWeb; accessed 1April 2016), and Wilson et al. (2013) scored it at the upper end of the medium vulnerability level. Data available on the life history and biology of A. ordinarium is restricted to the species description (Taylor, 1940), distribution (Shaffer, 1984; Anderson and Worthington, 1971), diet composition (Alvarado-Díaz et al., 2002), phylogeny (Weisrock et al., 2006) and the effect of habitat quality on diet diversity (Ruiz-Martínez et al., 2014). We did not find predation records on this species in the literature, and in this note we present information on a predation attack on an adult neotenic A. ordinarium by a Thamnophis cyrtopsis. On 13 July 2010 at 1300 h, while conducting an ecological study of A. -

Herpetological Information Service No
Type Descriptions and Type Publications OF HoBART M. Smith, 1933 through June 1999 Ernest A. Liner Houma, Louisiana smithsonian herpetological information service no. 127 2000 SMITHSONIAN HERPETOLOGICAL INFORMATION SERVICE The SHIS series publishes and distributes translations, bibliographies, indices, and similar items judged useful to individuals interested in the biology of amphibians and reptiles, but unlikely to be published in the normal technical journals. Single copies are distributed free to interested individuals. Libraries, herpetological associations, and research laboratories are invited to exchange their publications with the Division of Amphibians and Reptiles. We wish to encourage individuals to share their bibliographies, translations, etc. with other herpetologists through the SHIS series. If you have such items please contact George Zug for instructions on preparation and submission. Contributors receive 50 free copies. Please address all requests for copies and inquiries to George Zug, Division of Amphibians and Reptiles, National Museum of Natural History, Smithsonian Institution, Washington DC 20560 USA. Please include a self-addressed mailing label with requests. Introduction Hobart M. Smith is one of herpetology's most prolific autiiors. As of 30 June 1999, he authored or co-authored 1367 publications covering a range of scholarly and popular papers dealing with such diverse subjects as taxonomy, life history, geographical distribution, checklists, nomenclatural problems, bibliographies, herpetological coins, anatomy, comparative anatomy textbooks, pet books, book reviews, abstracts, encyclopedia entries, prefaces and forwords as well as updating volumes being repnnted. The checklists of the herpetofauna of Mexico authored with Dr. Edward H. Taylor are legendary as is the Synopsis of the Herpetofalhva of Mexico coauthored with his late wife, Rozella B. -

Intrageneric Relationships Among Colubrid Snakes of the Genus Geophis Wagler
MISCELLANEOUS PUBLICATIONS MUSEUM OF ZOOLOGY, UNIVERSITY OF MICHIGAN, NO. 131 Intrageneric Relationships Among Colubrid Snakes of the Genus Geophis Wagler BY FLOYD LESLIE DOWNS College of Wooster, Wooster, Ohio ANN ARBOR MUSEUM OF ZOOLOGY, UNIVERSITY OF MICHIGAN JULY 26, 1967 MISCELLANEOUS PUBLICATIONS MUSEUM OF ZOOLOGY, UNIVERSITY OF MICHIGAN The publications of the Museum of Zoology, University of Michigan, consist of two series-the Occasional Papers and the Miscellaneous Publications. Both series were founded by Dr. Bryant Walker, Mr. Bradshaw H. Swales, and Dr. W. W. Newcomb. The Occasional Papers, publication of which was begun in 1913, serve as a medium for original studies bascd principally upon the collections in the Museum. Thcy are issued separately. When a sufficient number of pages has been printed to make a volume, a title page, table of contents, and an index are supplied to libraries and indi- viduals on the mailing list for the series. The Miscellaneous Publications, which include papers on field and museum techniques, monographic studies, and other contributions not within the scope of the Occasional Papers, are published separtely. It is not intended that they be grouped into volumes. Each number has a title page and, when necessary, a table of contents. A complete list of publications on Birds, Fishes, Insects, Mammals, Mollusks, and Reptiles and Amphibians is available. Address inquiries to the Director, Museum of Zoology, Ann Arbor, Michigan. LISTOF MISCELLANEOUSPUBLICATIONS ON REPTILES AND AMPHIBIANS No. The amphibians and reptiles of the Sierra Nevada de Santa Marta, Colom- bia. By ALEXANDERG. RUTHVEN.(1922) 69 pp., 12 pls., 2 figs., 1 map .. -

M Ieiican%Usdum
M oxfitateSieiican%usdum PUBLISHED BY THE AMERICAN MUSEUM OF NATURAL HISTORY CENTRAL PARK WEST AT 79TH STREET, NEW YORK, N. Y. I0024 NUMBER 2 260 AUGUST i8, I 966 A New Species of Geophis (Serpentes, Colubridae) from the State of Colima, Mexico BY CHARLES M. BOGERT1 AND ANN P. PORTER2 While searching for spiders and scorpions in Mexico in 1965, Willis J. Gertsch and Richard Hastings obtained a few amphibians and reptiles, including a small snake that was found near the town of Tonila, Jalisco. The site where the snake was taken is, however, south of the Jalisco boundary, in Colima. This specimen proved to belong to the genus Geophis, but it also became apparent that it possessed an array of characters previously unreported for any species of the group. The snake evidently has affinities with species in the state of Michoacan. Nevertheless, the specimen from Colima differs in so many respects from those described that it is virtually certain that it represents an additional species. As a means of expressing our thanks to Dr. Gertsch for many useful specimens given to the Department over a period of years, as well as to credit him with the discovery of the species described below, we are naming it Geophis gertschi, new species HOLOTYPE: A.M.N.H. No. 94877, a male, presumably an adult, taken 2 miles south of Tonila, Jalisco, but in the state of Colima, at the northern extremity of the Distrito de Cuauhtemoc. The snake was found in moist soil beneath a rock by Willis J. Gertsch and Richard Hastings on August 28, 1965. -

Rare Snakes—Five New Species from Eastern Panama: Reviews Of
PUBLISHED BY THE AMERICAN MUSEUM OF NATURAL HISTORY CENTRAL PARK WEST AT 79TH STREET, NEW YORK, NY 10024 Number 3391, 47 pp., 20 ®gures, 2 maps, 3 tables February 19, 2003 Rare SnakesÐFive New Species from Eastern Panama: Reviews of Northern Atractus and Southern Geophis (Colubridae: Dipsadinae) CHARLES W. MYERS1 CONTENTS Abstract ....................................................................... 2 Resumen ....................................................................... 2 Introduction .................................................................... 3 Gorgas Memorial Laboratory and the Panama Snake Census ....................... 4 Methods of Study ............................................................. 5 Genus Atractus ................................................................. 8 Key to Panamanian Atractus ................................................... 9 Atractus clarki Dunn and Bailey .............................................. 10 Atractus darienensis, new species .............................................. 16 Atractus depressiocellus, new species .......................................... 20 Atractus hostilitractus, new species ............................................ 22 Atractus imperfectus, new species ............................................. 25 Genus Geophis ................................................................ 28 Key to Panamanian Geophis .................................................. 28 Records Excluded from Central Panama ........................................ 29 Geophis -

Geophis Nigroalbus Boulenger, 1908 (Serpentes, Dipsadidae), in the Magdalena Valley, Cordillera Oriental of Colombia
16 1 NOTES ON GEOGRAPHIC DISTRIBUTION Check List 16 (1): 115–119 https://doi.org/10.15560/16.1.115 First record of the goo-eater snake, Geophis nigroalbus Boulenger, 1908 (Serpentes, Dipsadidae), in the Magdalena Valley, Cordillera Oriental of Colombia Daniela García-Cobos1, Alejandro Corrales-García2, Gladys Cárdenas-Arévalo3, Azarys Paternina-Hernández3, Andrés Rymel Acosta-Galvis1 1 Subdirección de Investigaciones, Colecciones Biológicas, Instituto de Investigación de Recursos Biológicos Alexander von Humboldt, Carrera 8 # 15–08, Claustro de San Agustín, Villa de Leyva, Boyacá, Colombia. 2 Museo de Historia Natural ANDES, Departamento de Ciencias Biológicas, Facultad de Ciencias, Universidad de los Andes. Apartado 4976, Bogotá, Colombia. 3 Universidad Pedagógica y Tecnológica de Colombia, Avenida Central del Norte 39–115, Tunja, Boyacá, Colombia. Corresponding author: Daniela García-Cobos, [email protected] Abstract Geophis nigroalbus Boulenger, 1908 is a fossorial and secretive colubrid snake endemic to the western Andes of Colombia. Here we report this species for the first time in the Cordillera Oriental in Middle Magdalena Valley. We expand the known distribution of G. nigroalbus 183 km east of its original range. Keywords Boyacá, Colubridae, endemic, range extension. Academic editor: Ross MacCulloch | Received 24 October 2019 | Accepted 6 January 2020 | Published 7 February 2020 Citation: García-Cobos D, Corrales-García A, Cárdenas-Arévalo G, Paternina-Hernández A, Acosta-Galvis AR (2020) First record of the goo-eater snake, Geophis nigroalbus Boulenger, 1908 (Serpentes, Dipsadidae), in the Magdalena Valley, Cordillera Oriental of Colombia. Check List 16 (1): 115–119. https://doi.org/10.15560/16.1.115 Introduction region (Myers 2003; Wilson and Townsend 2007). -

Dipsadidae, Geophis) from Mexico 131 Doi: 10.3897/Zookeys.610.8605 RESEARCH ARTICLE Launched to Accelerate Biodiversity Research
A peer-reviewed open-access journal ZooKeys 610: 131–145 (2016)A new species of earth snake (Dipsadidae, Geophis) from Mexico 131 doi: 10.3897/zookeys.610.8605 RESEARCH ARTICLE http://zookeys.pensoft.net Launched to accelerate biodiversity research A new species of earth snake (Dipsadidae, Geophis) from Mexico Luis Canseco-Márquez1, Carlos J. Pavón-Vázquez1, Marco Antonio López-Luna2, Adrián Nieto-Montes de Oca1 1 Museo de Zoología and Departamento de Biología Evolutiva, Facultad de Ciencias, Universidad Nacio- nal Autónoma de México, Cd. Universitaria, Del. Coyoacán, Mexico 04510, Ciudad de México, Mexico 2 División Académica de Ciencias Biológicas, Universidad Juárez Autónoma de Tabasco, km 0.5 carretera Villahermosa-Cárdenas, entronque con Bosques de Saloya, Villahermosa, Tabasco, Mexico Corresponding author: Adrián Nieto-Montes de Oca ([email protected]) Academic editor: Z.T. Nagy | Received 23 March 2016 | Accepted 19 July 2016 | Published 11 August 2016 http://zoobank.org/1DFD1F68-B32A-41ED-8537-9B9F0B60329E Citation: Canseco-Márquez L, Pavón-Vázquez CJ, López-Luna MA, Nieto-Montes de Oca A (2016) A new species of earth snake (Dipsadidae, Geophis) from Mexico. ZooKeys 610: 131–145. doi: 10.3897/zookeys.610.8605 Abstract A new species of the Geophis dubius group is described from the mountains of the Sierra Zongolica in west-central Veracruz and the Sierra de Quimixtlán in central-east Puebla. The new species is most similar to G. duellmani and G. turbidus, which are endemic to the mountains of northern Oaxaca and the Sierra Madre Oriental of Puebla and Hidalgo, respectively. However, the new species differs from G. duellmani by the presence of postocular and supraocular scales and from G. -

Diversidad De Serpientes Y Estrategias Para Su Conservación En El Parque Nacional Natural Munchique, Departamento Del Cauca, Colombia
DIVERSIDAD DE SERPIENTES Y ESTRATEGIAS PARA SU CONSERVACIÓN EN EL PARQUE NACIONAL NATURAL MUNCHIQUE, DEPARTAMENTO DEL CAUCA, COLOMBIA 1 DIVERSIDAD DE SERPIENTES Y ESTRATEGIAS PARA SU CONSERVACIÓN EN EL PARQUE NACIONAL NATURAL MUNCHIQUE, DEPARTAMENTO DEL CAUCA, COLOMBIA Informe final presentado a Parques Nacionales Naturales de Colombia para dar cumplimiento a las obligaciones pertinentes establecidas en el permiso individual de recolección de especímenes de especies silvestres de la diversidad biológica con fines de investigación científica no comercial – Expediente PIDB DTPA No. 028 – 14; otorgado mediante resolución No. 187 de diciembre 17 de 2014. Por: 1 Luis Enrique Vera Pérez Investigador principal Y Jorge Alberto Zúñiga Baos Co-investigador 1 Documento de identidad: C.C. 1081400277 de La Plata Dirección: Carrera 8 # 2A – 12 La Plata, Huila Teléfono: 3113360894, 8370459 e-mail: [email protected] 2 CONTENIDO Pág. AGRADECIMIENTOS ………………………………………………………………………………………………...……... 6 RESUMEN ……………………………………………………………………………………………………………………….. 6 INTRODUCCIÓN ……………………………………………………...…………………………………………………….… 7 1. METODOLOGÍA ………………………………………………………………………………………………….………… 8 1.1 Área de estudio ………………….……………………………...……………………………………………………… 8 1.2 Educación ambiental ……………………………………........………………………………….………………. 10 1.3 Muestreos ………………………………………………………………………………………………………………. 10 1.4 Análisis de la información ………………………………………………………………………………………… 14 2. RESULTADOS ………………………………………………………………………………….…………………………. 15 2.1 Educación ambiental ………………………………………………………………….……………………………. -

Crotalus Tancitarensis. the Tancítaro Cross-Banded Mountain Rattlesnake
Crotalus tancitarensis. The Tancítaro cross-banded mountain rattlesnake is a small species (maximum recorded total length = 434 mm) known only from the upper elevations (3,220–3,225 m) of Cerro Tancítaro, the highest mountain in Michoacán, Mexico, where it inhabits pine-fir forest (Alvarado and Campbell 2004; Alvarado et al. 2007). Cerro Tancítaro lies in the western portion of the Transverse Volcanic Axis, which extends across Mexico from Jalisco to central Veracruz near the 20°N latitude. Its entire range is located within Parque Nacional Pico de Tancítaro (Campbell 2007), an area under threat from manmade fires, logging, avocado culture, and cattle raising. This attractive rattlesnake was described in 2004 by the senior author and Jonathan A. Campbell, and placed in the Crotalus intermedius group of Mexican montane rattlesnakes by Bryson et al. (2011). We calculated its EVS as 19, which is near the upper end of the high vulnerability category (see text for explanation), its IUCN status has been reported as Data Deficient (Campbell 2007), and this species is not listed by SEMARNAT. More information on the natural history and distribution of this species is available, however, which affects its conservation status (especially its IUCN status; Alvarado-Díaz et al. 2007). We consider C. tancitarensis one of the pre-eminent flagship reptile species for the state of Michoacán, and for Mexico in general. Photo by Javier Alvarado-Díaz. Amphib. Reptile Conserv. | http://amphibian-reptile-conservation.org 128 September 2013 | Volume 7 | Number 1 | e71 Copyright: © 2013 Alvarado-Díaz et al. This is an open-access article distributed under the terms of the Creative Commons Attribution–NonCommercial–NoDerivs 3.0 Unported License, which permits unrestricted use for Amphibian & Reptile Conservation 7(1): 128–170. -

Zootaxa: a Checklist and Key to the Snakes of the Genus Geophis
Zootaxa 1395: 1–31 (2007) ISSN 1175-5326 (print edition) www.mapress.com/zootaxa/ ZOOTAXA Copyright © 2007 · Magnolia Press ISSN 1175-5334 (online edition) A checklist and key to the snakes of the genus Geophis (Squamata: Colubridae: Dipsadinae), with commentary on distribution and conservation LARRY DAVID WILSON1 & JOSIAH H. TOWNSEND2 1Department of Biology, Miami-Dade College, Kendall Campus, Miami, Florida 33176-3393. 2School of Natural Resources and Environment, University of Florida, and Division of Herpetology, Florida Museum of Natural His- tory, Gainesville, Florida 32611-7800. Table of contents Abstract ...............................................................................................................................................................................2 Resumen ..............................................................................................................................................................................2 Introduction .........................................................................................................................................................................2 Acknowledgments ...............................................................................................................................................................3 Checklist ..............................................................................................................................................................................4 Geophis Wagler ...................................................................................................................................................................4 -

The HERPETOLOGICAL BULLETIN Number 115 – Spring 2011
The HERPETOLOGICAL BULLETIN Number 115 – Spring 2011 PUBLISHED BY THE BRITISH HERPETOLOGICAL SOCIETY THE HERPETOLOGICAL BULLETIN Contents RESEA R CH AR TICLES Notes on reproduction of Jackson’s chameleon Chamaeleo jacksonii (Squamata, Chaemaeleonidae), from Hawaii Stephen R. Goldberg and Fred Kraus................................. 1 Use of artificial wildlife ponds by reptiles in eastern Texas Cory K. Adams and Daniel Saenz.................................... 4 Assessment of an established population of atypical grass snakes Natrix natrix in the Aire Valley, UK Darryn J. Nash . 12 Pigmentation loss and regeneration in a captive wild-type axolotl, Ambystoma mexicanum James Barnett . 17 Notes on reptiles inhabiting a secondary, post development habitat, south Paphos, west Cyprus Frank D. Bowles ................................................ 19 Discovery of an extant population of the critically endangered treefrog Plectrohyla chrysopleura (Anura, Hylidae) in Refugio de Vida Silvestre Texiguat, Honduras Josiah H. Townsend, Larry David Wilson, Cesar A. Cerrato-M., Benjamin K. Atkinson, Luis A. Herrera-B. and Mayron M. Mejia.......... 22 Ecological data on road killed Amphisbaena alba Linnaeus, 1758 (Squamata, Amphisbaenidae) in southeast Brazil Carlos Henrique de Freitas and Atila Rodrigues de Araujo . 26 NATU R AL HISTO R Y NOTES Micrurus nigrocinctus (central American coral snake): Cannibalism Scott L. Travers, Stephen Doucette-Riise, Lenin A. Obando and Josiah H. Townsend .......................................... 31 Bothrops moojeni (Brazilian lancehead): Mating. Rogério L. Zacariotti, Taís F. Zimak and Rodrigo del Rio do Valle......... 33 Paleosuchus trigonatus (smooth-fronted caiman): Diet and movement. Sérgio A.A. Morato, Victor B.G.V. Batista and Anderson Paz ............. 34 Enyalius bibronii (NCN): Ectoparasitism Leonardo B. Ribeiro, Melissa Gogliath and Eliza M. Xavier Freire ........ 35 BOOK REVIEWS - Registered Charity No. -
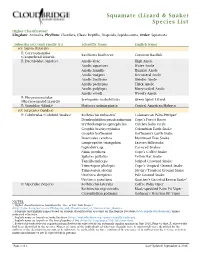
Squamate (Lizard & Snake) Species List
Squamate (Lizard & Snake) Species List Higher Classification1 Kingdom: Animalia, Phyllum: Chordata, Class: Reptilia, Diapsida, Lepidosauria, Order: Squamata Suborder (sO:) and Family (F:) Scientific Name2 English Name2 sO: Sauria (Lizards) F: Corytophanidae Basiliscus basiliscus Common Basilisk (Casquehead Lizards) F: Dactyloidae (Anoles) Anolis altae High Anole Anolis aquaticus Water Anole Anolis humilis Humble Anole Anolis insignis Decorated Anole Anolis limifrons Slender Anole Anolis pachypus Thick Anole Anolis polylepis Many-scaled Anole Anolis woodi Wood's Anole F: Phrynosomatidae Sceloporus malachiticus Green Spiny Lizard (Phrynosomatid Lizards) F: Scincidae (Skinks) Marisora unimarginata Central American Mabuya sO: Serpentes (Snakes) F: Colubridae (Colubrid Snakes) Bothriechis nubestris5 Talamancan Palm-Pitviper5 Dendrophidion paucicarinatum Cope's Forest Racer Erythrolamprus epinephelus Culebra boba verde Geophis brachycephalus Colombian Earth Snake Geophis hoffmanni Hoffmann's Earth Snake Imantodes cenchoa Blunthead Tree Snake Lampropeltis triangulum Eastern Milksnake Leptodeira sp. Cat-eyed Snakes Ninia psephota Cope's Coffee Snake Spilotes pullatus Yellow Rat Snake Tantilla ruficeps Striped Crowned Snake3 Trimetopon pliolepis Cope's Tropical Ground Snake Trimetopon slevini Slevin's Tropical Ground Snake Urotheca decipiens Pale Ground Snake Urotheca guentheri Gunther's Graceful Brown Snake4 F: Viperidae (Vipers) Bothriechis lateralis Coffee Palm Viper Bothriechis nigroviridis5 Black-speckled Palm Pit Viper5 Cerrophidion godmani Godman's Montane Pit Viper NOTES: 1, Higher classification as found on the Tree of Life Web Project (http://tolweb.org/accessory/Phylogeny_and_Classification_of_Amniotes?acc_id=462). 2, Scientific and English names based on current classifications as found on The Reptile Database (www.reptile-database.org), unless indicated otherwise. 3, English name as found on SnakeDatabase (http://snakedatabase.org/species/tantilla/ruficeps). 4, English name as found on The Encyclopedia of Life (http://eol.org/pages/1055095/overview).