Female and Hermaphrodite Flowers on a Chimeric Gynomonoecious
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Pinery Provincial Park Vascular Plant List Flowering Latin Name Common Name Community Date
Pinery Provincial Park Vascular Plant List Flowering Latin Name Common Name Community Date EQUISETACEAE HORSETAIL FAMILY Equisetum arvense L. Field Horsetail FF Equisetum fluviatile L. Water Horsetail LRB Equisetum hyemale L. ssp. affine (Engelm.) Stone Common Scouring-rush BS Equisetum laevigatum A. Braun Smooth Scouring-rush WM Equisetum variegatum Scheich. ex Fried. ssp. Small Horsetail LRB Variegatum DENNSTAEDIACEAE BRACKEN FAMILY Pteridium aquilinum (L.) Kuhn Bracken-Fern COF DRYOPTERIDACEAE TRUE FERN FAMILILY Athyrium filix-femina (L.) Roth ssp. angustum (Willd.) Northeastern Lady Fern FF Clausen Cystopteris bulbifera (L.) Bernh. Bulblet Fern FF Dryopteris carthusiana (Villars) H.P. Fuchs Spinulose Woodfern FF Matteuccia struthiopteris (L.) Tod. Ostrich Fern FF Onoclea sensibilis L. Sensitive Fern FF Polystichum acrostichoides (Michaux) Schott Christmas Fern FF ADDER’S-TONGUE- OPHIOGLOSSACEAE FERN FAMILY Botrychium virginianum (L.) Sw. Rattlesnake Fern FF FLOWERING FERN OSMUNDACEAE FAMILY Osmunda regalis L. Royal Fern WM POLYPODIACEAE POLYPODY FAMILY Polypodium virginianum L. Rock Polypody FF MAIDENHAIR FERN PTERIDACEAE FAMILY Adiantum pedatum L. ssp. pedatum Northern Maidenhair Fern FF THELYPTERIDACEAE MARSH FERN FAMILY Thelypteris palustris (Salisb.) Schott Marsh Fern WM LYCOPODIACEAE CLUB MOSS FAMILY Lycopodium lucidulum Michaux Shining Clubmoss OF Lycopodium tristachyum Pursh Ground-cedar COF SELAGINELLACEAE SPIKEMOSS FAMILY Selaginella apoda (L.) Fern. Spikemoss LRB CUPRESSACEAE CYPRESS FAMILY Juniperus communis L. Common Juniper Jun-E DS Juniperus virginiana L. Red Cedar Jun-E SD Thuja occidentalis L. White Cedar LRB PINACEAE PINE FAMILY Larix laricina (Duroi) K. Koch Tamarack Jun LRB Pinus banksiana Lambert Jack Pine COF Pinus resinosa Sol. ex Aiton Red Pine Jun-M CF Pinery Provincial Park Vascular Plant List 1 Pinery Provincial Park Vascular Plant List Flowering Latin Name Common Name Community Date Pinus strobus L. -

Diversity of Sexual Systems Within Different Lineages of the Genus Silene
Research Article Diversity of sexual systems within different lineages of the genus Silene Ine´s Casimiro-Soriguer1,2*, Maria L. Buide1 and Eduardo Narbona1 1 A´ rea de Bota´nica, Departamento de Biologı´a Molecular e Ingenierı´a Bioquı´mica, Universidad Pablo de Olavide, Ctra. de Utrera, km 1, 41013 Sevilla, Spain 2 A´ rea de Bota´nica, Departamento de Biologı´a Vegetal y Ecologı´a, Universidad de Sevilla, Avenida Reina Mercedes s/n, 41012 Sevilla, Spain Received: 16 December 2014; Accepted: 26 March 2015; Published: 10 April 2015 Associate Editor: Diana Wolf Citation: Casimiro-Soriguer I, Buide ML, Narbona E. 2015. Diversity of sexual systems within different lineages of the genus Silene. AoB PLANTS 7: plv037; doi:10.1093/aobpla/plv037 Abstract. Species and populations can be categorized by their sexual systems, depending on the spatial distribu- tion of female and male reproductive structures within and among plants. Although a high diversity of sexual systems exists in Silene, their relative frequency at the genus and infrageneric level is unknown. Here, we carried out an exten- sive literature search for direct or indirect descriptions of sexual systems in Silene species. We found descriptions of sexual systems for 98 Silene species, where 63 and 35 correspond to the phylogenetically supported subgenera Silene and Behenantha, respectively. Hermaphroditism was the commonest sexual system (58.2 %), followed by dioecy (14.3 %), gynodioecy (13.3 %) and gynodioecy–gynomonoecy (i.e. hermaphroditic, female and gynomonoecious plants coexisting in the same population; 12.2 %). The presence of these sexual systems in both subgenera suggests their multiple origins. -

Long-Read Transcriptome and Other Genomic Resources for the Angiosperm Silene Noctiflora
bioRxiv preprint doi: https://doi.org/10.1101/2020.08.09.243378; this version posted August 10, 2020. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available under aCC-BY-NC-ND 4.0 International license. Long-read transcriptome and other genomic resources for the angiosperm Silene noctiflora Alissa M. Williams,*,1 Michael W. Itgen,* Amanda K. Broz,* Olivia G. Carter,* Daniel B. Sloan* *Department of Biology, Colorado State University, Fort Collins, Colorado 80523 1Corresponding author: [email protected] bioRxiv preprint doi: https://doi.org/10.1101/2020.08.09.243378; this version posted August 10, 2020. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available under aCC-BY-NC-ND 4.0 International license. 1 Abstract 2 3 The angiosperm genus Silene is a model system for several traits of ecological and evolutionary 4 significance in plants, including breeding system and sex chromosome evolution, host-pathogen 5 interactions, invasive species biology, heavy metal tolerance, and cytonuclear interactions. 6 Despite its importance, genomic resources for this large genus of approximately 850 species are 7 scarce, with only one published whole-genome sequence (from the dioecious species S. latifolia). 8 Here, we provide genomic and transcriptomic resources for a hermaphroditic representative of 9 this genus (S. noctiflora), including a PacBio Iso-Seq transcriptome, which uses long-read, 10 single-molecule sequencing technology to analyze full-length mRNA transcripts and identify 11 paralogous genes and alternatively spliced genes. -

Investigating Past Range Dynamics for a Weed of Cultivation, Silene Vulgaris Megan E
Investigating past range dynamics for a weed of cultivation, Silene vulgaris Megan E. Sebasky1, Stephen R. Keller2 & Douglas R. Taylor1 1Department of Biology, University of Virginia, Charlottesville, Virginia 2Department of Plant Biology, University of Vermont, Burlington, Vermont Keywords Abstract Glacial refugia, last glacial maximum, MaxEnt, phylogeography, postglacial Since the last glacial maximum (LGM), many plant and animal taxa have expansion, Silene vulgaris, species distribution expanded their ranges by migration from glacial refugia. Weeds of cultivation model. may have followed this trend or spread globally following the expansion of agri- culture or ruderal habitats associated with human-mediated disturbance. We Correspondence tested whether the range expansion of the weed Silene vulgaris across Europe fit Megan E. Sebasky and Douglas R. Taylor, the classical model of postglacial expansion from southern refugia, or followed Department of Biology, University of Virginia, 485 McCormick Road, PO Box 400328, known routes of the expansion of human agricultural practices. We used spe- Charlottesville, VA 22904. cies distribution modeling to predict spatial patterns of postglacial expansion Tel: 434-982-5217; and contrasted these with the patterns of human agricultural expansion. A pop- Fax: 434-982-5626; ulation genetic analysis using microsatellite loci was then used to test which E-mails: [email protected] and scenario was better supported by spatial patterns of genetic diversity and struc- [email protected] ture. Genetic diversity was highest in southern Europe and declined with increasing latitude. Locations of ancestral demes from genetic cluster analysis Funding Information were consistent with areas of predicted refugia. Species distribution models National Science Foundation (Grant/Award Number: ‘MCB1022128’). -

Ethnobotanical Review of Wild Edible Plants in Spain
Blackwell Publishing LtdOxford, UKBOJBotanical Journal of the Linnean Society0024-4074The Linnean Society of London, 2006? 2006 View metadata, citation and similar papers1521 at core.ac.uk brought to you by CORE 2771 Original Article provided by Digital.CSIC EDIBLE WILD PLANTS IN SPAIN J. TARDÍO ET AL Botanical Journal of the Linnean Society, 2006, 152, 27–71. With 2 figures Ethnobotanical review of wild edible plants in Spain JAVIER TARDÍO1*, MANUEL PARDO-DE-SANTAYANA2† and RAMÓN MORALES2 1Instituto Madrileño de Investigación y Desarrollo Rural, Agrario y Alimentario (IMIDRA), Finca El Encín, Apdo. 127, E-28800 Alcalá de Henares, Madrid, Spain 2Real Jardín Botánico, CSIC, Plaza de Murillo 2, E-28014 Madrid, Spain Received October 2005; accepted for publication March 2006 This paper compiles and evaluates the ethnobotanical data currently available on wild plants traditionally used for human consumption in Spain. Forty-six ethnobotanical and ethnographical sources from Spain were reviewed, together with some original unpublished field data from several Spanish provinces. A total of 419 plant species belonging to 67 families was recorded. A list of species, plant parts used, localization and method of consumption, and harvesting time is presented. Of the seven different food categories considered, green vegetables were the largest group, followed by plants used to prepare beverages, wild fruits, and plants used for seasoning, sweets, preservatives, and other uses. Important species according to the number of reports include: Foeniculum vulgare, Rorippa nasturtium-aquaticum, Origanum vulgare, Rubus ulmifolius, Silene vulgaris, Asparagus acutifolius, and Scolymus hispanicus. We studied data on the botanical families to which the plants in the different categories belonged, over- lapping between groups and distribution of uses of the different species. -

Taxonomy and Conservation in Higher Plants and Bryophytes in the Mediterranean Area
Biodiversity Journal , 2015, 6 (1): 197–204 MONOGRAPH Taxonomy and conservation in Higher Plants and Bryophytes in the Mediterranean Area Gianniantonio Domina *, Giuseppe Bazan, Patrizia Campisi & Werner Greuter Orto botanico ed Herbarium Mediterraneum dell’Università di Palermo, via Lincoln 2, 90133 Palermo, Italy *Corresponding author ABSTRACT The Mediterranean Region is among the areas of the world richest in wild and cultivated taxa. Extinctions in the Mediterranean area are bound to have occurred in historical times but they are not documented. The probable and documented cases of plant extinction in specific areas within the Mediterranean are equivalent to 0.25% of total species-by-area records. Species with a large range are more prone to local population size fluctuations and eventual extinction than species with a reduced population. Small islands floras are more prone to extinction than those on large islands and on the mainland. Reliability of our data on Mediterranean plant ex - tinctions is poor. New emphasis on floristic research is needed to boost our deficient knowledge of the Mediterranean flora. A closer collaboration between scholars and amateurs can increase floristic knowledge and also help unravel taxonomic problems. KEY WORDS vascular plants; mosses; extinctions; nomenclature. Received 11.01.2015; accepted 02.03.2015; printed 30.03.2015 Proceedings of the 2nd International Congress “Speciation and Taxonomy”, May 16th-18th 2014, Cefalù-Castelbuono (Italy) INTRODUCTION et al., 2009) is an outstanding example. Global summaries for Europe have been presented by The Mediterranean Region is among the areas Sharrock & Jones (2009), Bilz et al. (2011), and of the world richest in wild and cultivated taxa. -

Title: Heavy Metals Bioaccumulation and Physiological Responses To
Title: Heavy metals bioaccumulation and physiological responses to heavy metal stress in populations of Silene vulgaris Moench (Garcke) from heavy metal contaminated sites Author: Aleksandra Nadgórska-Socha, Ryszard Ciepał, Marta Kandziora, Alina Kafel Citation style: Nadgórska-Socha Aleksandra, Ciepał Ryszard, Kandziora Marta, Kafel Alina (2009). Heavy metals bioaccumulation and physiological responses to heavy metal stress in populations of Silene vulgaris Moench (Garcke) from heavy metal contaminated sites. „Ecological Chemistry and Engineering A” (2009, no. 4, s. 389-397) ECOLOGICALCHEMISTRYANDENGINEERINGA Vol. 16, No. 4 2009 Aleksandra NADGÓRSKA-SOCHA1, Ryszard CIEPA£1, Marta KANDZIORA1 and Alina KAFEL2 HEAVY METALS BIOACCUMULATION AND PHYSIOLOGICAL RESPONSES TO HEAVY METAL STRESS IN POPULATIONS OF Silene vulgaris Moench (Garcke) FROM HEAVY METAL CONTAMINATED SITES BIOAKUMULACJA METALI CIʯKICH I ODPOWIED FIZJOLOGICZNA ROŒLIN Silene vulgaris Moench (Garcke) Z TERENÓW ZANIECZYSZCZONYCH METALAMI CIʯKIMI Abstract: In this study we have evaluated the heavy metals accumulation in the leaves of Silene vulgaris and its influence on glutathione, free amino acids and anthocyanins concentrations. Glutathione is a tripeptide (�-Glu-Cys-Gly) which is involved in many metabolic processes of a plant cell. It is related to the sequestration of xenobiotics and heavy metals and it is also an essential component of the antioxidant system, which keeps reactive oxygen species under control. The accumulation of anthocyanin pigments in the leaves can -

From Cacti to Carnivores: Improved Phylotranscriptomic Sampling And
Article Type: Special Issue Article RESEARCH ARTICLE INVITED SPECIAL ARTICLE For the Special Issue: Using and Navigating the Plant Tree of Life Short Title: Walker et al.—Phylotranscriptomic analysis of Caryophyllales From cacti to carnivores: Improved phylotranscriptomic sampling and hierarchical homology inference provide further insight into the evolution of Caryophyllales Joseph F. Walker1,13, Ya Yang2, Tao Feng3, Alfonso Timoneda3, Jessica Mikenas4,5, Vera Hutchison4, Caroline Edwards4, Ning Wang1, Sonia Ahluwalia1, Julia Olivieri4,6, Nathanael Walker-Hale7, Lucas C. Majure8, Raúl Puente8, Gudrun Kadereit9,10, Maximilian Lauterbach9,10, Urs Eggli11, Hilda Flores-Olvera12, Helga Ochoterena12, Samuel F. Brockington3, Michael J. Moore,4 and Stephen A. Smith1,13 Manuscript received 13 October 2017; revision accepted 4 January 2018. 1 Department of Ecology & Evolutionary Biology, University of Michigan, 830 North University Avenue, Ann Arbor, MI 48109-1048 USA 2 Department of Plant and Microbial Biology, University of Minnesota-Twin Cities, 1445 Gortner Avenue, St. Paul, MN 55108 USA 3 Department of Plant Sciences, University of Cambridge, Cambridge CB2 3EA, UK 4 Department of Biology, Oberlin College, Science Center K111, 119 Woodland Street, Oberlin, OH 44074-1097 USA 5 Current address: USGS Canyonlands Research Station, Southwest Biological Science Center, 2290 S West Resource Blvd, Moab, UT 84532 USA 6 Institute of Computational and Mathematical Engineering (ICME), Stanford University, 475 Author Manuscript Via Ortega, Suite B060, Stanford, CA, 94305-4042 USA This is the author manuscript accepted for publication and has undergone full peer review but has not been through the copyediting, typesetting, pagination and proofreading process, which may lead to differences between this version and the Version of Record. -

Chickpea 1 Chickpea
Chickpea 1 Chickpea Chickpea Varieties Left, Bengal (Indian); right, European Scientific classification Kingdom: Plantae (unranked): Angiosperms (unranked): Eudicots (unranked): Rosids Order: Fabales Family: Fabaceae Genus: Cicer Species: C. arietinum Binomial name Cicer arietinum L. The chickpea (Cicer arietinum) is a legume of the family Fabaceae, subfamily Faboideae. Its seeds are high in protein. It is one of the earliest cultivated legumes: 7,500-year-old remains have been found in the Middle East.[1] Other common names for the species include garbanzo bean, ceci bean, chana, sanagalu, Gonzo Bean and Bengal gram. Etymology The name "chickpea" traces back through the French chiche to cicer, Latin for ‘chickpea’ (from which the Roman cognomen Cicero was taken). The Oxford English Dictionary lists a 1548 citation that reads, "Cicer may be named in English Cich, or ciche pease, after the Frenche tonge." The dictionary cites "Chick-pea" in the mid-18th century; the original word in English taken directly from French was chich, found in print in English in 1388 and became obsolete in the 18th century. The word garbanzo came to English as "calavance" in the 17th century, from Old Spanish (perhaps influenced by Old Spanish garroba or algarroba), though it came to refer to a variety of other beans (cf. Calavance). The Portuguese (?) arvanço has suggested to some that the origin of the word garbanzo is in the Greek erebinthos.[2] But the Oxford English Dictionary notes that some scholars doubt this; it also mentions a possible origination in the word Chickpea 2 garbantzu, from Basque — a non-Indo-European tongue — in which it is a compound of garau, seed + antzu, dry. -
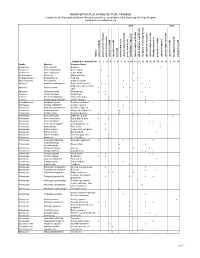
Preliminary Plant List
Growing Plant List of RoeJan Park, Hillsdale compiled by the Participatory Natural History Survey Group, an initiative of the Farmscape Ecology Program [email protected] 2009 2008 e e d i d i s - s - e n e e e W d a d d d W w i w i i i J e s o o s s s d - e g - - - d d w a o d W S a a S S E W a o o e f R t e e r t l E d d s w w t w w s w f E s E E a a a M m W d s o f e o o o o o e w m l N l e o N N r r r r r t r r r e r d r e o a t r e t t e e e e s o e e i a o a m r f d s f f s s o e g g g F g e / w S t w m f y o a e r e e d d r d d a e r d o o r r a e n h o m l l n l e n e a e i e e i o f f i o o h s w f e F r d f a h m h h f f h y i d a l d r f a d l l t l n n n n b p i p i a p d d d d d d d r r r r a n p a c b t y d n n n n n n i n e e e e f m w a i m W d l t t u l t t n a a a a a o a a a n t a b r l o l l l l l l r a s s s s o c f e W a h w W p o p p p a p l p i p a a a a w l l N u f e e H S w u u u e e b u s a S F u S p u Habitat # on Aerial Photo 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 20 15 5 18 19 Family Species Common Name Aceraceae Acer negundo Boxelder x Aceraceae Acer saccharinum Silver maple x Aceraceae Acer saccharum Sugar maple x Alismataceae Alisma sp. -

Scientific Name Symbol Common Name Rank Family Est. Occurrence Acer Negundo L
Non-Native Plant Species List – Alaska Center for Conservation Science Scientific Name Symbol Common Name Rank Family Est. Occurrence Acer negundo L. ACNE2 boxelder Aceraceae Adjacent Canada only Achillea filipendulina Lam. ACFI fernleaf yarrow Asteraceae 2005 Alaska Achillea ptarmica L. ACPT sneezeweed 46 Asteraceae 1913 Alaska Achnatherum hymenoides (Roem. & Schult.) Barkworth ACHY Indian ricegrass Poaceae Alaska (unconfirmed) Acorus calamus L. ACCA4 calamus Acoraceae 1968 Alaska Acroptilon repens (L.) DC. ACRE3 hardheads 66 Asteraceae Adjacent Canada only Aegopodium podagraria L. AEPO bishop's goutweed 57 Apiaceae 2006 Alaska Agropyron cristatum (L.) Gaertn. AGCR crested wheatgrass Poaceae 1939 Alaska Agropyron desertorum (Fisch. ex Link) J.A. Schult. AGDE2 desert wheatgrass Poaceae 1941 Alaska Agropyron fragile (Roth) P. Candargy AGFR Siberian wheatgrass Poaceae Adjacent Canada only Agrostemma githago L. AGGI common corncockle Caryophyllaceae 2006 Alaska Agrostis capillaris L. AGCA5 colonial bentgrass Poaceae 1941 Alaska Agrostis gigantea Roth AGGI2 redtop Poaceae 1902 Alaska Agrostis stolonifera L. AGST2 creeping bentgrass Poaceae 1918 Alaska Aira caryophyllea L. AICA silver hairgrass Poaceae Adjacent Canada only Alchemilla mollis (Buser) Rothm. ALMO12 lady's mantle 56 Rosaceae 2006 Alaska Alchemilla monticola Opiz ALMO4 hairy lady's mantle 56 Rosaceae 1979 Alaska Alliaria petiolata (M. Bieb.) Cavara & Grande ALPE4 garlic mustard 70 Brassicaceae 2002 Alaska Alnus glutinosa (L.) Gaertn. ALGL2 European alder 61 Betulaceae Alaska Watchlist Alopecurus arundinaceus Poir. ALAR creeping meadow foxtail Poaceae 2011 Alaska Alopecurus geniculatus L. ALGE2 water foxtail 49 Poaceae 1940 Alaska Alopecurus pratensis L. ALPR3 meadow foxtail 52 Poaceae 1934 Alaska Alyssum alyssoides (L.) L. ALAL3 alyssum Brassicaceae 1954 Alaska Amaranthus albus L. AMAL prostrate pigweed Amaranthaceae 2003 Alaska Amaranthus retroflexus L. -

Phylogenetic Relationships of Silene Sect. Melandrium and Allied Taxa
! "#! $ %&'(%)($ *++%,-,./,0 *+)%12.,.--0.1-,,. ! 3 4 3 3!!35.,6626 !" # $%&%&&'&'()&* * * + ,- . , / ",%&&',+ / *, " - 0 1 * $ 2 -," , 3)',4& , ,'5!6' 6774657 63, -* *8 0 1 " 9 * : * , * 8 ," . ; * : * , * * 0 1,- * * ,- " ,- ; : * . , < . ** * =>? . 6 +/ ,- ; , " * , ! , $ 8* " ,@ * * ,* ! * , - . * **, "# $= $ = =-"=- " "/" /+"%/+%/+%=> =? <-= @ %&' ( ) *+,-./# A" 8/ %&&' <== 37 63% 4 <='5!6' 6774657 63 ( ((( 6 &&!&30 (BB ,:,B C D ( ((( 6 &&!&31 Cover illustrations: Barry Breckling, Sara Gold, Kjell Lännerholm, Anja Rautenberg and Wiebke Rautenberg. List of Papers This thesis is based on the following papers, which are referred to in the text by their Roman numerals. I Rautenberg, A., Filatov, D., Svennblad, B., Heidari, N., Oxel- man, B. (2008) Conflicting phylogenetic signals in the SlX1/Y1 gene in Silene. BMC Evolutionary Biology, 8:299 II Rautenberg, A., Hathaway, L., Oxelman, B., Prentice, H. C. Phylogenetic relationships of Silene section Elisanthe (Caryo- phyllaceae) as inferred from chloroplast and nuclear DNA se- quences. Manuscript. III Rautenberg, A., Hathaway, L., Prentice, H. C., Oxelman, B. Phylogenetic relationships