Isopoda: Sphaeromatidae B
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Decreto N° (Pratica VDS018 21)
Decreto n° (pratica VDS018_21) DPR 357/97 art. 5. Regolamento recante attuazione della direttiva 92/43/CEE relativa alla conservazione degli habitat naturali e seminaturali, nonché della flora e della fauna selvatiche. Delibera di Giunta regionale n. 1323 del 11/07/2014. Indirizzi applicativi in materia di valutazione d’incidenza. Intervento: Programma delle immissioni a scopo di ripopolamento di Trota marmorata (Salmo marmoratus). Primo lotto 2021. VDS018_21. Proponente: ETPI - Ente tutela patrimonio ittico Comuni: Tramonti di sopra, Pontebba, Preone, Forni di Sopra, Forni Avoltri, Paularo, Forni di Sotto, Forni Avoltri, Osoppo, Nimis, Pulfero.Il Direttore del Servizio biodiversità Vista la direttiva del Consiglio 21 maggio 1992, n. 92/43/CEE relativa alla conservazione degli habitat naturali e seminaturali e della flora e della fauna selvatiche; Visto in particolare l’articolo 6, paragrafo 3 della suddetta direttiva 92/43/CEE, il quale prevede che qualsiasi piano o progetto non direttamente connesso e necessario alla gestione del sito ma che possa avere incidenze significative su tale sito, singolarmente o congiuntamente ad altri piani o progetti, deve essere sottoposto a valutazione d’incidenza, tenuto conto degli obiettivi di conservazione del medesimo; Visto il decreto del Presidente della Repubblica 8 settembre 1997, n. 357 regolamento recante attuazione della direttiva 92/43/CEE; Visto in particolare l’articolo 5 del suddetto DPR 357/1997, disciplinante la valutazione d’incidenza; Vista la delibera di Giunta regionale n. 1323 del 11/07/2014 recante indirizzi applicativi in materia di valutazione d’incidenza. Visto il decreto del Presidente della Regione n. 0277/Pres. del 27 agosto 2004 e successive modifiche e integrazioni recante il Regolamento di organizzazione dell’Amministrazione regionale e degli Enti regionali; Vista la deliberazione della Giunta regionale n. -

Drenchia, Grimacco, Prepotto, Pulfero, San Leonardo, San Pietro Al Natisone, Savogna, Stregna, Torreano
AZIENDA TERRITORIALE PER L’EDILIZIA RESIDENZIALE DI UDINE 33100 Udine, Via Sacile n. 15 – Tel. 0432 491111 – Fax 0432 546438 e-mail: [email protected] - web: udine.aterfvg.it Commissione per l’accertamento dei requisiti soggettivi di cui agli artt. 11 e 21, comma 6, della legge regionale 6 agosto 2019, n. 14 BANDO DI CONCORSO N. 4 / 2019 del 19 luglio 2019 per l'assegnazione in locazione di alloggi di edilizia residenziale pubblica sovvenzionata siti nel Comune o nei Comuni di DRENCHIA, GRIMACCO, PREPOTTO, PULFERO, SAN LEONARDO, SAN PIETRO AL NATISONE, SAVOGNA, STREGNA, TORREANO. GRADUATORIA DEFINITIVA L’assegnazione degli alloggi agli aventi diritto avverrà in base alla presente graduatoria, tenuto conto delle caratteristiche degli stessi alloggi e della composizione del nucleo familiare destinatario dell’assegnazione. La presente graduatoria è resa pubblica mediante affissione, per trenta giorni consecutivi, nella sede dell’ATER, in luogo aperto al pubblico, all’albo pretorio e nelle sedi di decentramento comunale del Comune o dei Comuni nei cui territori hanno sede gli alloggi oggetto del bando, nei siti web istituzionali della Regione, dell’ATER e dei Comuni medesimi. I termini minimi di pubblicazione previsti ai fini di legittimità sono esclusivamente quelli risultanti dal sito internet dell’ATER. A parità di punteggio, è stata considerata prioritaria la domanda presentata dal richiedente residente da più tempo in Regione e, in subordine, la domanda presentata dal richiedente già presente in graduatorie precedenti dello stesso Comune o comprensorio di Comuni, senza soluzione di continuità; al perdurare della parità, le domande sono state inserite in graduatoria previo sorteggio. Nel rispetto di quanto previsto dall’art. -

Comune Di Torreano
COMUNE DI TORREANO ESERCIZIO FINANZIARIO 2017 RELAZIONE DELL’ORGANO ESECUTIVO Art. 151, comma 6, D.Lgs. 267/2000 CONTO CONSUNTIVO ESERCIZIO FINANZIARIO 2017 RELAZIONE ILLUSTRATIVA DELLA GIUNTA COMUNALE L’art. 151 del D.Lgs. 267/2000, comma 6°, prescrive che al rendiconto dei Comuni sia allegata una relazione della Giunta sulla gestione che esprime le valutazioni di efficacia dell'azione condotta sulla base dei risultati conseguiti. Anche l’art. 231 del D.Lgs. 267/2000 prescrive: “La relazione sulla gestione è un documento illustrativo della gestione dell'ente, nonché dei fatti di rilievo verificatisi dopo la chiusura dell'esercizio, contiene ogni eventuale informazione utile ad una migliore comprensione dei dati contabili, ed è predisposto secondo le modalità previste dall'art. 11, comma 6, del decreto legislativo 23 giugno 2011, n. 118, e successive modificazioni”. La presente relazione è quindi redatta per soddisfare il precetto legislativo, per fornire dati di ragguaglio sulla produzione dei servizi pubblici e per consentire una idonea valutazione della realizzazione delle previsioni di bilancio. Questo Comune al 31.12.2017 conta n. 2129 abitanti residenti nel capoluogo e nelle frazioni di: Reant, Masarolis, Canalutto, Costa, Ronchis, Montina, Prestento e Togliano Il Comune di Torreano si caratterizza per le modeste dimensioni geografiche e demografiche, per la montuosità del territorio e per l'insediamento sullo stesso di un'economia in cui vi è una distribuzione piuttosto omogenea delle varie attività (agricola, industriale, artigianale e terziaria). Particolarmente conosciuta è l'attività della lavorazione della pietra Piasentina. Nel corso del 2017 il Comune di Torreano ha visto ulteriormente ridurre il personale in servizio a seguito della cessazione per quiescenza di un proprio dipendente. -
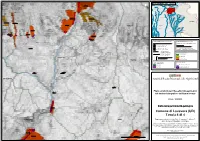
Comune Di Lusevera (UD) Tavola 4 Di 4
!< !< !< !< !< !< !< !< !< !< !< .! INAmQpezUzo ADRAM.!ENTO DEL.!LA TA.!VOLA Tolmezzo Moggio UdineseChiusaforte 0300510300A .! 0302041600 Cavazzo Carnico !< ± 1 2 0300510600 !< .! .!Gemona del Friuli !< Trasaghis .! 3 Lusevera 0302042500 !< 4 0302042400 .! !< Travesio .! Attimis 0302041300 .! !< !< .! Pulfero 0300510500 San Daniele del Friuli !<0300510500-CR 0300512300-CR .! .! 0302041500 Spilimbergo !< Povoletto .! !< 0302121700 Cividale del Friuli SLOVENIA 0302040000 !< !< .! 0302042400-CR !< UDINE 0302042500-CR 0302040100 0302039800 0302041200 !< !< 0300510200 !< 0300512300 .! !< 0300510600 !< Sedegliano 0302041400 0302039900 0302066800 0302121603 !< !< !< !< 0302040300 .! !< .! Codroipo San Giovanni al Natisone 0302040400 !< 0302040500 0302041000 0302058900 !< .! 0302040700 GORIZIA 0300510200 !< !< !<0300510400 0302127100 0302042900 !< .! 03020409000302041100 03020!<40200 !< !< !< !< 2 Palmanova .! 0302039600 Gradisca d'Isonzo !< 0302040800 !< 0302040600 0302039600 0302042900 !< !< 0302042600 !< !< 0300510400A .! .!Latisana San Canzian d'Isonzo 0302127300 0302121601 !< !< !< 0300510400 !< 0302127200 PIANO ASSETTO IDROGEOLOGICO P.A.I. ZONE DI ATTENZIONE GEOLOGICA !< !< Perimetrazione e classi di pericolosità geologica QUADRO CONOSCITIVO COMPLEMENTARE AL P.A.I. P1 - Pericolosità geologica moderata Banca dati I.F.F.I. - P2 - Pericolosità geologica media Inventario dei fenomeni franosi in Italia 0302350700 0302!<350700 P3 - Pericolosità geologica elevata !< Localizzazione dissesto franoso non delimitato P4 - Pericolosità geologica molto -

Cividale Del Friuli Moimacco, Povoletto Und Remanzacco Die
camper f Piazza della Resistenza P.tta i NOTRUFNUMMERN San Biagio u Castelmonte WIE UND WO m Slovenia Notarzt: 118 Colli orientali del Friuli: Landkarte und Führer und Landkarte Friuli: del orientali Colli e N 08 a Via P. Zorutti Carabinieri: 112 t i s o TARCENTO UND DIE TÄLER DES TORRE DES TÄLER DIE UND TARCENTO n e Staatspolizei: 113 Stadtpolizei: +39 0432 710220 CIVIDALE DEL FRIULI UND DIE VALLI DEL NATISONE DEL VALLI DIE UND FRIULI DEL CIVIDALE Borgo Brossana Gorizia - Aereoporto Feuerwehr: 115 04 Steuerfahndung: 117 Piazza Pannenhilfe: 803116 San Nicolò Via Scipione da Manzano Via Michele della Torre Via Montenero Via del Monastero Maggiore 07 P.tta Via del Monastero Maggiore Chiarottini ÖFFENTLICHE VERKEHRSMITTEL Die Valli Stretta T. Cerchiari Stretta S. M. di Corte Via Borgo di Ponte SAF-Busdienst Gebührenfreie Nummer: 800 915 303 03 www.saf.ud.it Piazza S. Giovanni del Natisone Pulfero P.tta Str.ta delle Giudaica 02 06 Zorutti FUC - Bahn Udine Cividale Piazza Duomo 05 ti C. Gallo NATUR UND TRADITIONEN HÖHLEN UND BÄREN ot tel +39 0432 731032 nd a C V Via Patriarcato 01 . i a B . www.ferrovieudinecividale.it S a G Via Cavour n a i Piazzetta L V Stretta J. Stellini Garibaldi a APT - Busdienst GÖRZ Gebührenfreie Nummer: 800 955 957 z rso Paolino D’Aquileia z C o a r Mitten im Grünen gelegen, ist es einer der Hauptorte der einer Verwaltungseinheit, der ursprünglich der langobardi- o www.aptgorizia.it Täler und bietet den Besuchern ein Netz von gut begehba- sche König vorstand, später der Patriarch von Aquileia und ino Die Valli del Natisone verbinden Cividale del Friuli mit dem Tal des Flusses Isonzo in Slowenien und nt le a Piazza V L a n a Paolo Diacono ren Wegen, um die Kunst- und Naturschätze zu entdecken. -

Curriculum Vitae
CURRICULUM VITAE INFORMAZIONI PERSONALI Nome Bonanni Marcella Data di nascita 15/09/1970 Qualifica Segretario comunale Amministrazione COMUNE DI TORREANO Incarico attuale Responsabile - Area Amministrativa Numero telefonico 0432712028 dell’ufficio Fax dell’ufficio 0432712345 E-mail istituzionale [email protected] TITOLI DI STUDIO E PROFESSIONALI ED ESPERIENZE LAVORATIVE Titolo di studio LAUREA IN GIURISPRUDENZA Altri titoli di studio e - professionali - corso di formazione teorico-pratico, presso il FORMEZ, Centro Formazione e Studi - con sede in Pozzuoli (Napoli), in materie giuridico-amministrative, finalizzato all’inserimento presso amministrazioni locali in qualità di funzionario - area vigilanza; - - corso-concorso organizzato dalla Scuola Superiore della Pubblica Amministrazione Locale (SSPAL), 18 mesi di lezioni più 6 mesi di tirocinio pratico presso i comuni di Sorrento (Na) , Buttrio (Ud), Trivignano Udinese (Ud), per l’accesso alla carriera di Segretario Comunale e provinciale,. L’accesso al corso-concorso è stato finalizzato alla formazione ed all’ingresso nella carriera di Segretario Comunale e Provinciale. Qualifica acquisita: Segretario Comunale di fascia C (fino ai 3.000 ab.). Dal dicembre del 2007 sono inquadrata nella fascia professionale B del CCNL di categoria (Segretario Generale) in seguito al superamento del corso SPES organizzato dalla Scuola Superiore dell’Amministrazione Locale (SSPAL); Esperienze professionali - (incarichi ricoperti) Dal 23 febbraio 2004 al 19 febbraio 2006: Segretario Comunale titolare del comune di Pulfero (UD); - COMUNE DI PULFERO - dal 20 febbraio 2006 al 31 ottobre 2008: Segretario Comunale titolare della sede convenzionata di Segreteria Pulfero-Montenars (UD); - COMUNE DI PULFERO - dal 01.11.2008 al 31.08.2009: Segretario Comunale della sede di segreteria convenzionata Pulfero (UD) – Savogna 1 CURRICULUM VITAE (UD) – Stregna (UD). -

Prefettura – UTG Di Trieste Albo Segretari Comunali E Provinciali Friuli Venezia Giulia Il Prefetto Della Provincia Di Trieste
Prefettura – UTG di Trieste Albo Segretari Comunali e Provinciali Friuli Venezia Giulia Trieste, data del protocollo TRASMESSO: Ai Sig.ri Sindaci dei comuni di PULFERO SAN LEONARDO Alla dott.ssa Eva STANIG c/o il comune di San Leonardo OGGETTO: incarico alla dott.ssa Eva Stanig della reggenza a scavalco presso la segreteria del comune di Pulfero dal 1° maggio 2020 al 30 giugno 2020. Il Prefetto della provincia di Trieste quale Prefetto del capoluogo della regione Friuli Venezia Giulia ai sensi dell’art. 7, comma 31-ter del D.L. n. 78/2010 convertito in legge n. 122/2010, e dell’art. 1 del decreto 23.5.2012 del Ministro dell’Interno di concerto con il Ministro dell’Economia e delle Finanze PREMESSO: - che la segreteria del comune di Pulfero, fascia di popolazione sino a 3.000 abitanti, ex classe IV, è vacante; - che il sindaco del comune di Pulfero, con nota prot. n. 1867 del 29.4.2020, ha chiesto di incaricare della reggenza a scavalco la dott.ssa Eva Stanig, titolare della segreteria convenzionata San Leonardo - Porpetto, dal 1° maggio 2020 al 30 giugno 2020; - che per lo svolgimento di detto incarico sono state acquisite l’autorizzazione del sindaco del comune di San Leonardo e l’accettazione del segretario; VISTA la Circolare del Ministero dell’Interno, Albo Segretari, prot. n. 23581 del 2013; VISTA la nota del Ministero dell’Interno, Albo Segretari acquisita al prot. n. 14102 del 19.3.2020, avente ad oggetto “Reggenze/Supplenze a scavalco dei segretari comunali e provinciali. Chiarimenti”, cui si rinvia per le valutazioni di competenza delle singole amministrazioni locali; VISTE le delibere n. -

COMUNE Di DRENCHIA Provincia Di Udine Piano Di Razionalizzazione
COMUNE di DRENCHIA Provincia di Udine Piano di razionalizzazione delle società partecipate (art. 20 comma 2 D.lgs. 175/2016) Allegato alla Deliberazione Consiliare n. ______ TITOLO I - Introduzione generale 1. Premessa Dopo il “Piano Cottarelli”, con il quale l’allora commissario straordinario alla spending review auspicava la drastica riduzione delle società partecipate da circa 8.000 a circa 1.000, la legge di stabilità per il 2015 (legge 190/2014) ha imposto agli enti locali l’avvio un “processo di razionalizzazione” che possa produrre risultati già entro fine 2015. Il comma 611 della legge 190/2014 dispone che, allo scopo di assicurare il “coordinamento della finanza pubblica, il contenimento della spesa, il buon andamento dell'azione amministrativa e la tutela della concorrenza e del mercato”, gli enti locali devono avviare un “processo di razionalizzazione” delle società e delle partecipazioni, dirette e indirette, che permetta di conseguirne una riduzione entro il 31 dicembre 2015. Lo stesso comma 611 indica i criteri generali cui si deve ispirare il “processo di razionalizzazione”: a) eliminare le società e le partecipazioni non indispensabili al perseguimento delle finalità istituzionali, anche mediante liquidazioni o cessioni; b) sopprimere le società che risultino composte da soli amministratori o da un numero di amministratori superiore a quello dei dipendenti; c) eliminare le partecipazioni in società che svolgono attività analoghe o similari a quelle svolte da altre società partecipate o da enti pubblici strumentali, anche mediante operazioni di fusione o di internalizzazione delle funzioni; d) aggregare società di servizi pubblici locali di rilevanza economica; e) contenere i costi di funzionamento, anche mediante la riorganizzazione degli organi amministrativi e di controllo e delle strutture aziendali, ovvero riducendo le relative remunerazioni. -

Regione Friuli-Venezia Giulia
Regione Friuli-Venezia Giulia Direzione regionale dell'Ambiente MONITORAGGIO INQUINAMENTO DA METALLI IN AREE DELLA REGIONE CONTAMINATE DA RILASCI DI SOSTANZE PERICOLOSE MEDIANTE L'USO DI LICHENI COME BIOACCUMULATORI 1 I N T R O D U Z I O N E Nel Marzo del 1997 ha avuto inizio la fase attuativa di uno studio, commissionato dalla Regione Friuli-Venezia Giulia al Dipartimento di Biologia dell' Universit à di Trieste, mirante al biomonitoraggio di metalli in traccia tramite licheni in aree contaminate da rilasci incidentali o dolosi di sostanze pericolose. Il lavoro, come previsto dalla apposita convenzione, è sta to suddiviso nelle seguenti fasi operative: PRIMA FASE: in questa fase la Convenzione prevedeva il campionamento di licheni epifiti e la successiva analisi di 10 metalli in 114 stazioni di rilevamento, la cui localizzazione sul territorio regionale è stata regolarmente fornita dalla Regione all' Università. Il lavoro di campionamento è stato effettuato a partire dal 10 Maggio 1997, e si è protratto sino al 12 Giugno. Sono state visitate accuratamente tutte le 114 stazioni specificate dalla Regione. E' stato possibile reperire campioni di licheni atti all'analisi spettrofotometrica in 99 stazioni. Il materiale è stato preparato e sottoposto ad analisi spettrofotometrica. SECONDA FASE: la seconda fase, della prevista durata di due mesi, prevedeva un' elaborazione numerica ed un riporto cartografico preliminari relativi ai dati acquisiti nell'ambito della prima fase. Nella seconda relazione intermedia sono stati forniti i valori relativi ai metalli, e le relative carte. TERZA FASE: la terza fase, della durata prevista di due mesi, prevedeva il campionamento in ulteriori 40 stazioni (aggiuntive rispetto alle 114 previste per la prima fase) per giungere al totale previsto di 154 stazioni. -

Second Report Submitted by Italy Pursuant to Article 25, Paragraph 1 of the Framework Convention for the Protection of National Minorities
Strasbourg, 14 May 2004 ACFC/SR/II(2004)006 SECOND REPORT SUBMITTED BY ITALY PURSUANT TO ARTICLE 25, PARAGRAPH 1 OF THE FRAMEWORK CONVENTION FOR THE PROTECTION OF NATIONAL MINORITIES (received on 14 May 2004) MINISTRY OF THE INTERIOR DEPARTMENT FOR CIVIL LIBERTIES AND IMMIGRATION CENTRAL DIRECTORATE FOR CIVIL RIGHTS, CITIZENSHIP AND MINORITIES HISTORICAL AND NEW MINORITIES UNIT FRAMEWORK CONVENTION FOR THE PROTECTION OF NATIONAL MINORITIES II IMPLEMENTATION REPORT - Rome, February 2004 – 2 Table of contents Foreword p.4 Introduction – Part I p.6 Sections referring to the specific requests p.8 - Part II p.9 - Questionnaire - Part III p.10 Projects originating from Law No. 482/99 p.12 Monitoring p.14 Appropriately identified territorial areas p.16 List of conferences and seminars p.18 The communities of Roma, Sinti and Travellers p.20 Publications and promotional activities p.28 European Charter for Regional or Minority Languages p.30 Regional laws p.32 Initiatives in the education sector p.34 Law No. 38/2001 on the Slovenian minority p.40 Judicial procedures and minorities p.42 Database p.44 Appendix I p.49 - Appropriately identified territorial areas p.49 3 FOREWORD 4 Foreword Data and information set out in this second Report testify to the considerable effort made by Italy as regards the protection of minorities. The text is supplemented with fuller and greater details in the Appendix. The Report has been prepared by the Ministry of the Interior – Department for Civil Liberties and Immigration - Central Directorate for Civil Rights, Citizenship and Minorities – Historical and new minorities Unit When the Report was drawn up it was also considered appropriate to seek the opinion of CONFEMILI (National Federative Committee of Linguistic Minorities in Italy). -

Pedemontana Colline Pianura
Schede climatiche del Friuli Venezia Giulia SCHEDA CLIMATICA N. 22 osservabile: vento zona: COLLINE, PEDEMONTANA E PIANURA comuni di: Arba, Aviano *, Budoia *, Caneva di Sacile*, Cavasso Nuovo, Fanna, Maniago *, Montereale val Cellina *, Pinzano al Tagliamento, Polcenigo*, Sequals, Vajont, Arzene, Azzano Decimo, Brugnera, Chions, Cordenons, Cordovado, Fiume Veneto, Fontanafredda, Morsano al Tagliamento, Pasiano di Pordenone, Porcia, Pordenone, Prata di Pordenone, Pravisdomini, Roveredo in Piano, Sacile, San Giorgio della Richinvelda, San Martino al Tagliamento, San Quirino, San Vito al Tagliamento, Sesto al Reghena, Spilimbergo, Valvasone, Vivaro, Zoppola (PN); Aiello, Aquileia, Artegna, Attimis *, Bagnaria Arsa, Basiliano, Bertiolo, Bicinicco, Buia, Buttrio, Camino al Tagliamento, Campoformido, Campolongo Tapogliano, Carlino, Cassacco, Castions di Strada, Cervignano, Chiopris, Cividale del Friuli, Codroipo, Colloredo di Montalbano, Corno di Rosazzo, Coseano, Dignano, Faedis *, Fagagna, Fiumicello**, Flaibano, Gonars, Latisana, Lestizza, Magnano, Majano, Manzano, Martignacco, Mereto di Tomba, Moimacco, Mortegliano, Moruzzo, Muzzana, Nimis*, Osoppo, Pagnacco, Palazzolo dello Stella, Palmanova, Pasian di Prato, Pocenia, Porcia di Udine, Porpetto, Povoletto, Pozzuolo del Friuli, Pradamano, Precenicco, Premariacco, Prepotto *, Ragogna, Reana del Roiale, Remanzacco, Rive d'Arcano, Rivignano Teor, Ronchis, Ruda, San Daniele del Friuli, San Giorgio di Nogaro, San Giovanni al Natisone, San Vito al Torre, San Vito di Fagagna, Santa Maria la Longa, Sedegliano, Talmassons, Tarcento *, Tavagnacco, Terzo d'Aquileia, Torreano *, Torviscosa, Treppo Grande, Tricesimo, Trivignano Udinese, Udine, Varmo, Villa Vicentina, Visco (UD); Capriva, Cormons, Dolegna del Collio, Farra d'Isonzo, Fogliano, Gorizia, Gradisca, Mariano, Medea, Moraro, Mossa, Romans, Ronchi dei Legionari, San Canzian d'Isonzo, San Lorenzo Isontino, San Pier d'Isonzo, Savogna d'Isonzo, San Floriano del Collio, Staranzano**, Turriaco, Villesse (GO). -

Natisone Nasce Dal Desiderio Di Natisone Riunire, in Un Unico Ambito Di Tutela Ambientale E Di Un Fiume in Festa
Il progetto per l’istituzione del Parco fluviale tran- sfrontaliero del Natisone nasce dal desiderio di Natisone riunire, in un unico ambito di tutela ambientale e di un fiume in festa sviluppo ecosostenibile, i territori che si dispiegano 151812 lungo l’asta del fiume Natisone e che comprendono i Si ringraziano Siti di Interesse Comunitario di Taipana e Gran Mon- te, Magredi di Firmano, confluenza Natisone-Torre Le Amministrazioni comunali di inseriti nel progetto europeo “Natura 2000”. Taipana, Caporetto/Kobarid, Pulfero, San Pietro al Natisone, www.sincromia.it Il Comitato Promotore ha redatto, e presentato ai Cividale del Friuli, Premariacco, Manzano, San Giovanni al Natisone, Trivignano Udinese, Comuni del Natisone e alla Regione Friuli Venezia Chiopris-Viscone per l’avvio dell’iter istitutivo del Parco Giulia, un documento che illustra le grandi ricchezze e la collaborazione alla realizzazione di FESTINPARCO naturalistiche, storiche, culturali, artistiche, agrarie ed enogastronomiche del territorio del Natisone e Gruppo F.A.I. Cividale del Friuli propone un piano di valorizzazione sinergica che sia Club UNESCO di Udine piattaforma di rilancio economico. Il Comitato Ipplis da ritrovare Le Pro Loco: Alcuni passi fondamentali sono stati compiuti nel 2015: Amici di Prossenicco - il Comitato promotore è diventato Associazione Pulfero Parco del Natisone Ponteacco - le Amministrazioni comunali di Taipana, Caporet- Facciamo Quadrato Trivignano Udinese to, Pulfero, San Pietro al Natisone, Cividale del Friuli, Premariacco, Manzano, San Giovanni al Natisone, Le Associazioni: Trivignano Udinese e Chiopris-Viscone, che, aderen- Il Villaggio degli Orsi di Pulfero Natisone in bici di Cividale del Friuli do al progetto per la realizzazione del Parco, hanno Circolo Culturale Ricreativo di Trivignano Udinese sottoscritto una convenzione con la Regione per la condivisione di tutti i dati concernenti il fiume.