FAU Institutional Repository
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

(Gastropoda: Littorinidae) in the Temperate Southern Hemisphere: the Genera Nodilittorina, Austrolittorina and Afrolittorina
© Copyright Australian Museum, 2004 Records of the Australian Museum (2004) Vol. 56: 75–122. ISSN 0067-1975 The Subfamily Littorininae (Gastropoda: Littorinidae) in the Temperate Southern Hemisphere: The Genera Nodilittorina, Austrolittorina and Afrolittorina DAVID G. REID* AND SUZANNE T. WILLIAMS Department of Zoology, The Natural History Museum, London SW7 5BD, United Kingdom [email protected] · [email protected] ABSTRACT. The littorinine gastropods of the temperate southern continents were formerly classified together with tropical species in the large genus Nodilittorina. Recently, molecular data have shown that they belong in three distinct genera, Austrolittorina, Afrolittorina and Nodilittorina, whereas the tropical species are members of a fourth genus, Echinolittorina. Austrolittorina contains 5 species: A. unifasciata in Australia, A. antipodum and A. cincta in New Zealand, and A. fernandezensis and A. araucana in western South America. Afrolittorina contains 4 species: A. africana and A. knysnaensis in southern Africa, and A. praetermissa and A. acutispira in Australia. Nodilittorina is monotypic, containing only the Australian N. pyramidalis. This paper presents the first detailed morphological descriptions of the African and Australasian species of these three southern genera (the eastern Pacific species have been described elsewhere). The species-level taxonomy of several of these has been confused in the past; Afrolittorina africana and A. knysnaensis are here distinguished as separate taxa; Austrolittorina antipodum is a distinct species and not a subspecies of A. unifasciata; Nodilittorina pyramidalis is separated from the tropical Echinolittorina trochoides with similar shell characters. In addition to descriptions of shells, radulae and reproductive anatomy, distribution maps are given, and the ecological literature reviewed. -

The Systematics and Ecology of the Mangrove-Dwelling Littoraria Species (Gastropoda: Littorinidae) in the Indo-Pacific
ResearchOnline@JCU This file is part of the following reference: Reid, David Gordon (1984) The systematics and ecology of the mangrove-dwelling Littoraria species (Gastropoda: Littorinidae) in the Indo-Pacific. PhD thesis, James Cook University. Access to this file is available from: http://eprints.jcu.edu.au/24120/ The author has certified to JCU that they have made a reasonable effort to gain permission and acknowledge the owner of any third party copyright material included in this document. If you believe that this is not the case, please contact [email protected] and quote http://eprints.jcu.edu.au/24120/ THE SYSTEMATICS AND ECOLOGY OF THE MANGROVE-DWELLING LITTORARIA SPECIES (GASTROPODA: LITTORINIDAE) IN THE INDO-PACIFIC VOLUME I Thesis submitted by David Gordon REID MA (Cantab.) in May 1984 . for the Degree of Doctor of Philosophy in the Department of Zoology at James Cook University of North Queensland STATEMENT ON ACCESS I, the undersigned, the author of this thesis, understand that the following restriction placed by me on access to this thesis will not extend beyond three years from the date on which the thesis is submitted to the University. I wish to place restriction on access to this thesis as follows: Access not to be permitted for a period of 3 years. After this period has elapsed I understand that James Cook. University of North Queensland will make it available for use within the University Library and, by microfilm or other photographic means, allow access to users in other approved libraries. All uses consulting this thesis will have to sign the following statement: 'In consulting this thesis I agree not to copy or closely paraphrase it in whole or in part without the written consent of the author; and to make proper written acknowledgement for any assistance which I have obtained from it.' David G. -

Marine Biological Investigations in the Bahamas 23. Description of the Littoral Zonation at Nine Bahamian Rocky-Shore Localities
Marine biological investigations in the Bahamas 23. Description of the littoral zonation at nine Bahamian rocky-shore localities Hans Brattström Brattström H. 1999. Description of the littoral zonation at nine Bahamian rocky-shore localities. SARSIA Sarsia 84:319-365. Nine typical, more or less vertical, rocky-shore localities were visited in April-May 1967, and one of these was revisited in March 1968. The topographies of the stations are described and illustrated, and the vertical zonation of their flora and fauna is documented. On the whole the flora and fauna at the stations investigated was fairly poor. At least 37 species of algae and 85 animal species were recorded, but only 15 algae and 71 animals could be identified to species. Most of the algae were too undeveloped or grazed down to permit a reliable identification. Only half as many species were found at these nine stations as were found on three almost horizontal beach-rock stations described in an earlier article. Whereas many of the latter lived in a system of cavities below the rocks, all species found on the nine vertical stations were exposed to waves and the hot midday sun. The common yellow, black, grey, and white zones typical of the Bahamas, were often well-devel- oped. Some algae and many of the animals formed distinct belts within these zones, the higher up the more exposed the rocks were. Though tides and exposure presumably are responsible for the general zonation pattern, it is likely that in the cirripeds and vermetids the way of food-intake determines their upper limit. -

Malacofauna Marina Del Parque Nacional “Los Caimanes”, Villa Clara, Cuba
Tesis de Diploma Malacofauna Marina del Parque Nacional “Los Caimanes”, Villa Clara, Cuba. Autora: Liliana Olga Quesada Pérez Junio, 2011 Universidad Central “Marta Abreu” de Las Villas Facultad Ciencias Agropecuaria TESIS DE DIPLOMA Malacofauna marina del Parque Nacional “Los Caimanes”, Villa Clara, Cuba. Autora: Liliana Olga Quesada Pérez Tutor: M. C. Ángel Quirós Espinosa Investigador Auxiliar y Profesor Auxiliar [email protected] Centro de Estudios y Servicios Ambientales, CITMA-Villa Clara Carretera Central 716, Santa Clara Consultante: Dr.C. José Espinosa Sáez Investigador Titular Instituto de Oceanología Junio, 2011 Pensamiento “La diferencia entre una mala observación y una buena, es que la primera es errónea y la segunda es incompleta.” Van Hise Dedicatoria Dedicatoria: A mis padres, a Yandy y a mi familia: por las innumerables razones que me dan para vivir, y por ser fuente de inspiración para mis metas. Agradecimientos Agradecimientos: Muchos son los que de alguna forma contribuyeron a la realización de este trabajo, todos saben cuánto les agradezco: Primero quiero agradecer a mis padres, que aunque no estén presentes sé que de una forma u otra siempre estuvieron allí para darme todo su amor y apoyo. A mi familia en general: a mi abuela, hermano, a mis tíos por toda su ayuda y comprensión. A Yandy y a su familia que han estado allí frente a mis dificultades. Agradecer a mi tutor el M.Sc. Ángel Quirós, a mi consultante el Dr.C. José Espinosa y a la Dra.C. María Elena, por su dedicación para el logro de esta tesis. A mis compañeros de grupo por estos cinco años que hemos compartidos juntos, que para mí fueron inolvidables. -

ABSTRACT Title of Dissertation: PATTERNS IN
ABSTRACT Title of Dissertation: PATTERNS IN DIVERSITY AND DISTRIBUTION OF BENTHIC MOLLUSCS ALONG A DEPTH GRADIENT IN THE BAHAMAS Michael Joseph Dowgiallo, Doctor of Philosophy, 2004 Dissertation directed by: Professor Marjorie L. Reaka-Kudla Department of Biology, UMCP Species richness and abundance of benthic bivalve and gastropod molluscs was determined over a depth gradient of 5 - 244 m at Lee Stocking Island, Bahamas by deploying replicate benthic collectors at five sites at 5 m, 14 m, 46 m, 153 m, and 244 m for six months beginning in December 1993. A total of 773 individual molluscs comprising at least 72 taxa were retrieved from the collectors. Analysis of the molluscan fauna that colonized the collectors showed overwhelmingly higher abundance and diversity at the 5 m, 14 m, and 46 m sites as compared to the deeper sites at 153 m and 244 m. Irradiance, temperature, and habitat heterogeneity all declined with depth, coincident with declines in the abundance and diversity of the molluscs. Herbivorous modes of feeding predominated (52%) and carnivorous modes of feeding were common (44%) over the range of depths studied at Lee Stocking Island, but mode of feeding did not change significantly over depth. One bivalve and one gastropod species showed a significant decline in body size with increasing depth. Analysis of data for 960 species of gastropod molluscs from the Western Atlantic Gastropod Database of the Academy of Natural Sciences (ANS) that have ranges including the Bahamas showed a positive correlation between body size of species of gastropods and their geographic ranges. There was also a positive correlation between depth range and the size of the geographic range. -

Resistant Pseudosuccinea Columella Snails to Fasciola Hepatica (Trematoda) Infection in Cuba : Ecological, Molecular and Phenotypical Aspects Annia Alba Menendez
Comparative biology of susceptible and naturally- resistant Pseudosuccinea columella snails to Fasciola hepatica (Trematoda) infection in Cuba : ecological, molecular and phenotypical aspects Annia Alba Menendez To cite this version: Annia Alba Menendez. Comparative biology of susceptible and naturally- resistant Pseudosuccinea columella snails to Fasciola hepatica (Trematoda) infection in Cuba : ecological, molecular and phe- notypical aspects. Parasitology. Université de Perpignan; Instituto Pedro Kouri (La Havane, Cuba), 2018. English. NNT : 2018PERP0055. tel-02133876 HAL Id: tel-02133876 https://tel.archives-ouvertes.fr/tel-02133876 Submitted on 20 May 2019 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. Délivré par UNIVERSITE DE PERPIGNAN VIA DOMITIA En co-tutelle avec Instituto “Pedro Kourí” de Medicina Tropical Préparée au sein de l’ED305 Energie Environnement Et des unités de recherche : IHPE UMR 5244 / Laboratorio de Malacología Spécialité : Biologie Présentée par Annia ALBA MENENDEZ Comparative biology of susceptible and naturally- resistant Pseudosuccinea columella snails to Fasciola hepatica (Trematoda) infection in Cuba: ecological, molecular and phenotypical aspects Soutenue le 12 décembre 2018 devant le jury composé de Mme. Christine COUSTAU, Rapporteur Directeur de Recherche CNRS, INRA Sophia Antipolis M. Philippe JARNE, Rapporteur Directeur de recherche CNRS, CEFE, Montpellier Mme. -

Guide to Theecological Systemsof Puerto Rico
United States Department of Agriculture Guide to the Forest Service Ecological Systems International Institute of Tropical Forestry of Puerto Rico General Technical Report IITF-GTR-35 June 2009 Gary L. Miller and Ariel E. Lugo The Forest Service of the U.S. Department of Agriculture is dedicated to the principle of multiple use management of the Nation’s forest resources for sustained yields of wood, water, forage, wildlife, and recreation. Through forestry research, cooperation with the States and private forest owners, and management of the National Forests and national grasslands, it strives—as directed by Congress—to provide increasingly greater service to a growing Nation. The U.S. Department of Agriculture (USDA) prohibits discrimination in all its programs and activities on the basis of race, color, national origin, age, disability, and where applicable sex, marital status, familial status, parental status, religion, sexual orientation genetic information, political beliefs, reprisal, or because all or part of an individual’s income is derived from any public assistance program. (Not all prohibited bases apply to all programs.) Persons with disabilities who require alternative means for communication of program information (Braille, large print, audiotape, etc.) should contact USDA’s TARGET Center at (202) 720-2600 (voice and TDD).To file a complaint of discrimination, write USDA, Director, Office of Civil Rights, 1400 Independence Avenue, S.W. Washington, DC 20250-9410 or call (800) 795-3272 (voice) or (202) 720-6382 (TDD). USDA is an equal opportunity provider and employer. Authors Gary L. Miller is a professor, University of North Carolina, Environmental Studies, One University Heights, Asheville, NC 28804-3299. -

UC Davis UC Davis Previously Published Works
UC Davis UC Davis Previously Published Works Title Molluscan marginalia: Serration at the lip edge in gastropods Permalink https://escholarship.org/uc/item/2mx5c6w9 Journal Journal of Molluscan Studies, 80(3) ISSN 0260-1230 Author Vermeij, GJ Publication Date 2014 DOI 10.1093/mollus/eyu020 Peer reviewed eScholarship.org Powered by the California Digital Library University of California Journal of The Malacological Society of London Molluscan Studies Journal of Molluscan Studies (2014) 80: 326–336. doi:10.1093/mollus/eyu020 Advance Access publication date: 16 April 2014 Molluscan marginalia: serration at the lip edge in gastropods Geerat J. Vermeij Geology Department, University of California, One Shields Avenue, Davis, CA 95616, USA Correspondence: G.J. Vermeij; e-mail: [email protected] Downloaded from (Received 5 September 2013; accepted 10 February 2014) ABSTRACT The shells of many marine gastropods have ventrally directed serrations (serial projections) at the edge http://mollus.oxfordjournals.org/ of the adult outer lip. These poorly studied projections arise as extensions either of external spiral cords or of interspaces between cords. This paper describes taxonomic, phylogenetic, architectural and func- tional aspects of serrations. Cord-associated serrations occur in cerithiids, strombids, the personid Distorsio anus, ocenebrine muricids and some cancellariids. Interspace-associated serrations are phylo- genetically much more widespread, and occur in at least 16 family-level groups. The nature of serration may be taxonomically informative in some fissurellids, littorinids, strombids and costellariids, among other groups. Serrated outer lips occur only in gastropods in which the apex points more backward than upward, but the presence of serrations is not a necessary byproduct of the formation of spiral sculp- tural elements. -

228100 N, Tomando El Camino De Los Conucos Hasta El Entronque
228100 N, tomando el camino de los Conucos hasta el entronque de Zapato en 101700 E, 231400 N, siguiendo por el camino de Zapato hasta el final en 100700 E, 231600 N, girando al N hasta el estero de Santa Catalina por el cual sale al mar en 100300 E, 234950 N, se continúa por la línea de costa hasta el nacimiento del estero Los Morros en 096800 E, 234900 N, el cual se dirige hasta el punto N de Barra Sorda en 094300 E, 235400 N, por la orilla NO de esta barra en su límite con los manglares hasta cruzar el camino de Faro Roncali al estero Palmarito en 091900 E, 231900 N, en dirección S a 150 m del terraplén del Sur hasta el Pantano de Caleta Larga en 092800 E, 229950 N, donde bordea el Pantano hasta 092600 E, 229850 N, siguiendo la línea recta virtual hasta 090800 E, 228750 N, punto inicial de este derrotero. • A los efectos de controlar adecuadamente las acciones que puedan repercutir negativamente sobre esta área protegida se establece una Zona de Amortiguamiento que comprende los 500 metros a partir del límite externo del área y que se indica en el Anexo Cartográfico. Anexo 3 Listado de especies de la flora Flora terrestre Flora marina División RHODOPHYTA Orden CORALLINALES Familia CORALLINACEA 1. Amphiroa beauvoisii Lamouroux 2. Amphiroa fragilissima (Linnaeus) Lamouroux 3. Amphiroa rigida Lamouroux 4. Haliptilon cubense (Montagne ex Kützing) Garbary et Johansen 5. Haliptilon subulatum (Ellis et Solander) Johansen 6. Hydrolithon farinosum (Lamouroux) Penrose et Chamberlain 7. Jania adhaerens Lamouroux 8. -
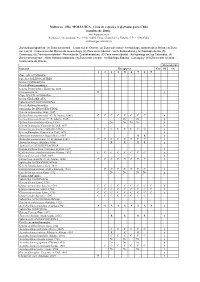
Moluscos - Filo MOLLUSCA
Moluscos - Filo MOLLUSCA. Lista de especies registradas para Cuba (octubre de 2006). José Espinosa Sáez Instituto de Oceanología, Ave 1ª No. 18406, Playa, Ciudad de La Habana, C.P. 11200, Cuba [email protected] Zonas biogeográficas: (1) Zona suroriental – Costa sur de Oriente, (2) Zona surcentral - Archipiélago Jardines de la Reina, (3) Zona sur central - Costa al sur del Macizo de Guamuhaya, (4) Zona suroccidental - Golfo de Batabanó y Archipiélago de los, (5) Canarreos, (6) Zona suroccidental - Península de Guanahacabibes, (7) Zona noroccidental - Archipiélago de Los Colorados, (8) Zona noroccidental - Norte Habana-Matanzas, (9) Zona norte-central - Archipiélago Sabana - Camagüey, (10) Zona norte-oriental - Costa norte de Oriente Abreviaturas Especies Bioegiones Cu Pl Oc 1 2 3 4 5 6 7 8 9 Clase APLACOPHORA Subclase SOLENOGASTRES Orden CAVIBELONIA Familia Proneomeniidae Género Proneomenia Hubrecht, 1880 Proneomenia sp . R x Clase POLYPLACOPHORA Orden NEOLORICATA Suborden ISCHNOCHITONINA Familia Ischnochitonidae Subfamilia ISCHNOCHITONINAE Género Ischnochiton Gray, 1847 Ischnochiton erythronotus (C. B. Adams, 1845) C C C C C C C C x Ischnochiton papillosus (C. B. Adams, 1845) Nc Nc x Ischnochiton striolatus (Gray, 1828) Nc Nc Nc Nc x Género Ischnoplax Carpenter in Dall, 1879 x Ischnoplax pectinatus (Sowerby, 1832) C C C C C C C C x Género Stenoplax Carpenter in Dall, 1879 x Stenoplax bahamensis Kaas y Belle, 1987 R R x Stenoplax purpurascens (C. B. Adams, 1845) C C C C C C C C x Stenoplax boogii (Haddon, 1886) R R R R x Subfamilia CALLISTOPLACINAE Género Callistochiton Carpenter in Dall, 1879 x Callistochiton shuttleworthianus Pilsbry, 1893 C C C C C C C C x Género Ceratozona Dall, 1882 x Ceratozona squalida (C. -

Temperature and Relative Humidity Effects on Water Loss and Hemolymph Osmolality of Littoraria Angulifera (Lamarck, 1822)
University of Texas Rio Grande Valley ScholarWorks @ UTRGV UTB/UTPA Electronic Theses and Dissertations Legacy Institution Collections 4-2014 Temperature and relative humidity effects on water loss and hemolymph osmolality of Littoraria angulifera (Lamarck, 1822) Phillip J. Rose The University of Texas Rio Grande Valley Follow this and additional works at: https://scholarworks.utrgv.edu/leg_etd Part of the Animal Sciences Commons, Environmental Sciences Commons, and the Oceanography and Atmospheric Sciences and Meteorology Commons Recommended Citation Rose, Phillip J., "Temperature and relative humidity effects on water loss and hemolymph osmolality of Littoraria angulifera (Lamarck, 1822)" (2014). UTB/UTPA Electronic Theses and Dissertations. 39. https://scholarworks.utrgv.edu/leg_etd/39 This Thesis is brought to you for free and open access by the Legacy Institution Collections at ScholarWorks @ UTRGV. It has been accepted for inclusion in UTB/UTPA Electronic Theses and Dissertations by an authorized administrator of ScholarWorks @ UTRGV. For more information, please contact [email protected], [email protected]. Temperature and Relative Humidity Effects on Water Loss and Hemolymph Osmolality of Littoraria angulifera (Lamarck, 1822) A Thesis Presented to the Faculty of the College of Science, Mathematics and Technology University of Texas at Brownsville In Partial Fulfillment of the Requirements for the Degree Master of Science In the field of Biology by Phillip J. Rose April 2014 Copyright By Phillip J. Rose April 2014 Acknowledgements I would like to acknowledge and especially thank the many people who assisted and/or contributed to this project in some way, shape, or form…………..and there were many! First, I would like to say a big thank you to the thesis committee comprised of Dr. -

Gastropod Molluscs of the Southern Area Of
PAIDEIA XXI Vol. 10, Nº 2, Lima, julio-diciembre 2020, pp. 289-310 ISSN Versión Impresa: 2221-7770; ISSN Versión Electrónica: 2519-5700 http://revistas.urp.edu.pe/index.php/Paideia ORIGINAL ARTICLE / ARTÍCULO ORIGINAL GASTROPOD MOLLUSCS OF THE SOUTHERN AREA OF CIENFUEGOS, FROM THE BEACH RANCHO LUNA TO THE MOUTH OF THE ARIMAO RIVER, CUBA MOLUSCOS GASTRÓPODOS DE LA ZONA SUR DE CIENFUEGOS, DESDE PLAYA RANCHO LUNA HASTA LA DESEMBOCADURA DEL RÍO ARIMAO, CUBA Oneida Calzadilla-Milian1*; Rafael Armiñana-García2,*; José Alexis Sarría- Martínez1; Rigoberto Fimia-Duarte3; Jose Iannacone4,5; Raiden Grandía- Guzmán6 & Yolepsy Castillo-Fleites2 1* Universidad de Cienfuegos «Carlos Rafael Rodríguez», Cienfuegos, Cuba. E-mail: ocalzadilla@ ucf.edu.cu; [email protected] 2 Universidad Central «Marta Abreu» de Las Villas, Villa Clara, Cuba. E-mail: rarminana@uclv. cu / ycfl [email protected] 3 Facultad de Tecnología de la Salud y Enfermería (FTSE). Universidad de Ciencias Médicas de Villa Clara (UCM-VC), Cuba. E-mail: rigoberto.fi [email protected] 4 Laboratorio de Ecología y Biodiversidad Animal (LEBA). Facultad de Ciencias Naturales y Matemáticas (FCNNM). Universidad Nacional Federico Villarreal (UNFV). Lima, Perú. 5 Facultad de Ciencias Biológicas. Universidad Ricardo Palma (URP). Lima, Perú. E-mail: [email protected] 6 Centro Nacional para la Producción de Animales de Laboratorio (CENPALAB), La Habana, Cuba. E-mail: [email protected] * Author for correspondence: [email protected] ABSTRACT The research presented shows a malacological survey of Cienfuegos' southern area, from “Rancho Luna” beach to the mouth of the “Arimao River”. The malacological studies ranged from January 2018 to December of the same year.