Jurassic-Cretaceous Transition on the Getic Carbonate Platform (Southern Carpathians, Romania): Benthic Foraminifera and Algae
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

94366148005.Pdf
Boletín de la Sociedad Geológica Mexicana ISSN: 1405-3322 Sociedad Geológica Mexicana, A.C. Omaña, Lourdes; López-Doncel, Rubén; Torres, José Ramón; Alencaster, Gloria; López-Caballero, Iriliana Mid-late Cenomanian larger benthic foraminifers from the El Abra Formation W Valles-San Luis Potosi Platform, central-eastern Mexico: Taxonomy, biostratigraphy and paleoenvironmental implications Boletín de la Sociedad Geológica Mexicana, vol. 71, no. 3, 2019, September-December, pp. 691-725 Sociedad Geológica Mexicana, A.C. DOI: https://doi.org/10.18268/BSGM2019v71n3a5 Available in: https://www.redalyc.org/articulo.oa?id=94366148005 How to cite Complete issue Scientific Information System Redalyc More information about this article Network of Scientific Journals from Latin America and the Caribbean, Spain and Journal's webpage in redalyc.org Portugal Project academic non-profit, developed under the open access initiative Boletín de la Sociedad Geológica Mexicana / 2019 / 691 Mid–late Cenomanian larger benthic foraminifers from the El Abra Formation W Valles-San Luis Potosi Platform, central–eastern Mexico: Taxonomy, biostratigraphy and paleoenvironmental implications Lourdes Omaña, Rubén López-Doncel, José Ramón Torres, Gloria Alencaster, Iriliana López-Caballero ABSTRACT Lourdes Omaña ABSTRACT RESUMEN Gloria Alencaster lomanya@geología.unam.mx The Valles–San Luis Potosí Platform is La Plataforma Valles–San Luis Potosí es parte de Departamento de Paleontologia, Instituto de part of an extensive carbonate platform un sistema de plataformas extenso que se encon- Geologia, Universidad Nacional Autonoma system that rimmed the ancestral Gulf traba bordeando el antiguo Golfo de México en el de Mexico, Ciudad Universitaria, Coyoacan, of Mexico during the middle (Albian) Albiano hasta el Cretácico Superior. -
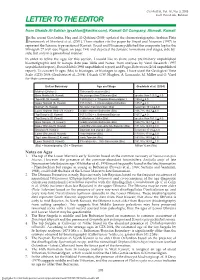
Letter to the Editor
GeoArabia, Vol. 10, No. 3, 2005 Gulf PetroLink, Bahrain LETTER TO THE EDITOR from Ghaida Al-Sahlan ([email protected]), Kuwait Oil Company, Ahmadi, Kuwait n the recent GeoArabia, Haq and Al-Qahtani (2005) updated the chronostratigraphic Arabian Plate Iframework of Sharland et al. (2001). These studies cite the paper by Yousif and Nouman (1997) to represent the Jurassic type section of Kuwait. Yousif and Nouman published the composite log for the Minagish-27 well (see Figure on page 194) and depicted the Jurassic formations and stages, side-by- side, but only in a generalized manner. In order to refine the ages for this section, I would like to share some preliminary unpublished biostratigraphic and Sr isotope data (see Table and Notes) from analyses by Varol Research (1997 unpublished report), ExxonMobil (1998 unpublished report) and Fugro-Robertson (2004 unpublished report). To convert Sr ages (Ma) to biostages, or biostages to ages, I have used the Geological Time Scale (GTS) 2004 (Gradstein et al., 2004). I thank G.W. Hughes, A. Lomando, M. Miller and O. Varol for their comments. Unit or Boundary Age and Stage Gradstein et al. (2004) Makhul (Offshore) Tithonian-Berriasian (Bio) Base Makhul (N. Kuwait) No younger than Tithonian (Bio) greater than 145.5 + 4.0 Top Hith (W. Kuwait) 150.0 (Sr) = c. Tithonian/Kimmeridgian ? 150.8 + 4.0 Upper Najmah (S. Kuwait) 155.0 (Sr) = c. Kimmeridgian/Oxfordian 155.7 + 4.0 Najmah (N. Kuwait) No older than Oxfordian (Bio) less than 161.2 + 4.0 Lower Najmah Shale (N. Kuwait) middle and late Bathonian (Bio) 166.7 to 164.7 + 4.0 Top Sargelu (S. -

The Late Jurassic Tithonian, a Greenhouse Phase in the Middle Jurassic–Early Cretaceous ‘Cool’ Mode: Evidence from the Cyclic Adriatic Platform, Croatia
Sedimentology (2007) 54, 317–337 doi: 10.1111/j.1365-3091.2006.00837.x The Late Jurassic Tithonian, a greenhouse phase in the Middle Jurassic–Early Cretaceous ‘cool’ mode: evidence from the cyclic Adriatic Platform, Croatia ANTUN HUSINEC* and J. FRED READ *Croatian Geological Survey, Sachsova 2, HR-10000 Zagreb, Croatia Department of Geosciences, Virginia Tech, 4044 Derring Hall, Blacksburg, VA 24061, USA (E-mail: [email protected]) ABSTRACT Well-exposed Mesozoic sections of the Bahama-like Adriatic Platform along the Dalmatian coast (southern Croatia) reveal the detailed stacking patterns of cyclic facies within the rapidly subsiding Late Jurassic (Tithonian) shallow platform-interior (over 750 m thick, ca 5–6 Myr duration). Facies within parasequences include dasyclad-oncoid mudstone-wackestone-floatstone and skeletal-peloid wackestone-packstone (shallow lagoon), intraclast-peloid packstone and grainstone (shoal), radial-ooid grainstone (hypersaline shallow subtidal/intertidal shoals and ponds), lime mudstone (restricted lagoon), fenestral carbonates and microbial laminites (tidal flat). Parasequences in the overall transgressive Lower Tithonian sections are 1– 4Æ5 m thick, and dominated by subtidal facies, some of which are capped by very shallow-water grainstone-packstone or restricted lime mudstone; laminated tidal caps become common only towards the interior of the platform. Parasequences in the regressive Upper Tithonian are dominated by peritidal facies with distinctive basal oolite units and well-developed laminate caps. Maximum water depths of facies within parasequences (estimated from stratigraphic distance of the facies to the base of the tidal flat units capping parasequences) were generally <4 m, and facies show strongly overlapping depth ranges suggesting facies mosaics. Parasequences were formed by precessional (20 kyr) orbital forcing and form parasequence sets of 100 and 400 kyr eccentricity bundles. -

Kimmeridgian (Late Jurassic) Cold-Water Idoceratids (Ammonoidea) from Southern Coahuila, Northeastern Mexico, Associated with Boreal Bivalves and Belemnites
REVISTA MEXICANA DE CIENCIAS GEOLÓGICAS Kimmeridgian cold-water idoceratids associated with Boreal bivalvesv. 32, núm. and 1, 2015, belemnites p. 11-20 Kimmeridgian (Late Jurassic) cold-water idoceratids (Ammonoidea) from southern Coahuila, northeastern Mexico, associated with Boreal bivalves and belemnites Patrick Zell* and Wolfgang Stinnesbeck Institute for Earth Sciences, Heidelberg University, Im Neuenheimer Feld 234, 69120 Heidelberg, Germany. *[email protected] ABSTRACT et al., 2001; Chumakov et al., 2014) was followed by a cool period during the late Oxfordian-early Kimmeridgian (e.g., Jenkyns et al., Here we present two early Kimmeridgian faunal assemblages 2002; Weissert and Erba, 2004) and a long-term gradual warming composed of the ammonite Idoceras (Idoceras pinonense n. sp. and trend towards the Jurassic-Cretaceous boundary (e.g., Abbink et al., I. inflatum Burckhardt, 1906), Boreal belemnites Cylindroteuthis 2001; Lécuyer et al., 2003; Gröcke et al., 2003; Zakharov et al., 2014). cuspidata Sachs and Nalnjaeva, 1964 and Cylindroteuthis ex. gr. Palynological data suggest that the latest Jurassic was also marked by jacutica Sachs and Nalnjaeva, 1964, as well as the Boreal bivalve Buchia significant fluctuations in paleotemperature and climate (e.g., Abbink concentrica (J. de C. Sowerby, 1827). The assemblages were discovered et al., 2001). in inner- to outer shelf sediments of the lower La Casita Formation Upper Jurassic-Lower Cretaceous marine associations contain- at Puerto Piñones, southern Coahuila, and suggest that some taxa of ing both Tethyan and Boreal elements [e.g. ammonites, belemnites Idoceras inhabited cold-water environments. (Cylindroteuthis) and bivalves (Buchia)], were described from numer- ous localities of the Western Cordillera belt from Alaska to California Key words: La Casita Formation, Kimmeridgian, idoceratid ammonites, (e.g., Jeletzky, 1965), while Boreal (Buchia) and even southern high Boreal bivalves, Boreal belemnites. -

Upper Bajocian– Callovian) of the Polish Jura Chain and Holy Cross Mountains (South-Central Poland)
1661-8726/07/010153-12 Swiss j. geosci. 100 (2007) 153–164 DOI 10.1007/s00015-007-1207-3 Birkhäuser Verlag, Basel, 2007 A diverse crinoid fauna from the Middle Jurassic (Upper Bajocian– Callovian) of the Polish Jura Chain and Holy Cross Mountains (south-central Poland) MARIUSZ A. SALAMON*& MICHA¸ ZATO¡ Key words: crinoids, Middle Jurassic, Poland, palaeobiogeography, taphonomy, epibiontism ABSTRACT ZUSAMMENFASSUNG A systematic account of a diverse crinoid fauna from the Middle Jurassic Aus mitteljurassischen (Bajocian–Callovian) Sedimenten des südlichen (Upper Bajocian–Callovian) of the Polish Jura Chain and Holy Cross Moun- Zentralpolens (Krakow–Cz´stochowa Hochland und Heilig-Kreuz Gebirge) tains (south-central Poland) is presented. The description is supplemented wird eine diverse Crinoidenfauna systematisch beschrieben und stratigra- with a list of all crinoid species found hitherto in the Tatra Mountains and the phisch eingestuft. Die Beschreibung wird durch eine Zusammenstellung sämt- Pieniny Klippen Belt (Poland), which were a part of the northern margin of licher Crinoiden-Spezies ergänzt, die bislang im Tatra-Gebirge und im Pieniny the Tethys during Middle Jurassic time. Balanocrinus hessi seems to be en- Klippen-Gürtel gefunden wurden. Beide Regionen waren während des Mitt- demic and established its own population in the epicontinental sea. Other leren Jura Teil des Nordrandes der Tethys. Balanocrinus hessi bildete eigen- stalked crinoids entered from the Tethys through the East-Carpathian Gate or ständige Populationen in diesem epikontinentalen Meeresbereich und scheint from a westerly way, and constitute a typical Mediterranean fauna. Stemless endemisch gewesen zu sein. Andere gestielte Crinoiden drangen aus der forms are regarded to be unsuccessful immigrants. -

High European Sauropod Dinosaur Diversity During Jurassic–Cretaceous Transition in Riodeva (Teruel, Spain)
CORE Metadata, citation and similar papers at core.ac.uk Provided by RERO DOC Digital Library [Palaeontology, Vol. 52, Part 5, 2009, pp. 1009–1027] HIGH EUROPEAN SAUROPOD DINOSAUR DIVERSITY DURING JURASSIC–CRETACEOUS TRANSITION IN RIODEVA (TERUEL, SPAIN) by RAFAEL ROYO-TORRES*, ALBERTO COBOS*, LUIS LUQUE*, AINARA ABERASTURI*, , EDUARDO ESPI´LEZ*, IGNACIO FIERRO*, ANA GONZA´ LEZ*, LUIS MAMPEL* and LUIS ALCALA´ * *Fundacio´n Conjunto Paleontolo´gico de Teruel-Dino´polis. Avda. Sagunto s ⁄ n. E-44002 Teruel, Spain; e-mail: [email protected] Escuela Taller de Restauracio´n Paleontolo´gica II del Gobierno de Arago´n. Avda. Sagunto s ⁄ n. E-44002 Teruel, Spain Typescript received 13 December 2007; accepted in revised form 3 November 2008 Abstract: Up to now, more than 40 dinosaur sites have (CPT-1074) referring to the Diplodocidae clade. New been found in the latest Jurassic – earliest Cretaceous remains from the RD-28, RD-41 and RD-43 sites, of the sedimentary outcrops (Villar del Arzobispo Formation) of same age, among which there are caudal vertebrae, are Riodeva (Iberian Range, Spain). Those already excavated, assigned to Macronaria. New sauropod footprints from the as well as other findings, provide a large and diverse Villar del Arzobispo Formation complete the extraordinary number of sauropod remains, suggesting a great diversity sauropod record coming to light in the area. The inclusion for this group in the Iberian Peninsula during this time. of other sauropods from different contemporaneous expo- Vertebrae and ischial remains from Riodevan site RD-13 sures in Teruel within the Turiasauria clade adds new evi- are assigned to Turiasaurus riodevensis (a species described dence of a great diversity of sauropods in Iberia during in RD-10, Barrihonda site), which is part of the the Jurassic–Cretaceous transition. -

7. Tithonian Benthic Foraminifers from Hole 901A1
Whitmarsh, R.B., Sawyer, D.S., Klaus, A., and Masson, D.G. (Eds.), 1996 Proceedings of the Ocean Drilling Program, Scientific Results, Vol. 149 7. TITHONIAN BENTHIC FORAMINIFERS FROM HOLE 901A1 Eric S. Collins,2 Wolfgang Kuhnt,3 David B. Scott2 ABSTRACT Dark gray laminated silty claystones (Unit II) drilled at Site 901 contain Tithonian benthic foraminifer assemblages that indicate a neritic depositional environment and probably dysaerobic bottom-water conditions. Three benthic foraminifer zones are distinguished within Unit II. The upper part of the unit is dominated by Spirillina polygyrata, contains Globospirillina spp. (Samples 149-901A-3R-1, 10-12 cm, to 149-901A-3R-1, 75-77 cm) and is interpreted as late Tithonian. Samples 149-901A- 3R-1, 87-89 cm, to 149-901A-6R-1, 74-76 cm, contain Epistomina uhligi and Lingulina franconica and are probably early Tithonian. The early Tithonian Neobulimina atlantica Zone is characterized by the occurrence of the zonal marker and Epis- tomina uhligi and reaches from Sample 149-901A-6R-1, 128-130 cm, to the base of the drilled-sequence. The sediments and benthic foraminiferal assemblage characteristics of the Tithonian-aged sequence in Hole 901A are unknown elsewhere in the Atlantic and may represent deposition in a marginal shelf basin with increased terrigenous and organic flux. INTRODUCTION the sediments, the samples were only wet-sieved through a 63-um (#230 mesh) screen and the residue was dried. Foraminifer residues Upper Jurassic sediments have been found at DSDP Sites 4,5,99, were examined using a binocular microscope with at least ×40 mag- 100, 105, 111, 367, 391, 401, 416, 534, 544-547 and ODP Site 639 nification. -

E. Geochemistry
APPENDIX E – GEOCHEMISTRY APPENDIX E - GEOCHEMISTRY TABLE OF CONTENTS APPENDIX E – GEOCHEMISTRY .............................................................................. APPENDIX E.3 LIST OF FIGURES Figure A.E. 1. Eel Formation source rock characteristics, offshore Ireland. ......................................................... Appendix E.7 Figure A.E. 2 Rainbow Claystone Formation source rock characteristics, offshore Ireland. ................................. Appendix E.8 Figure A.E. 3 Sybil Formation source rock characteristics, offshore Ireland. ....................................................... Appendix E.9 Figure A.E. 4 Galley Formation source rock characteristics, offshore Ireland. ................................................... Appendix E.10 Figure A.E. 5 Kestrel Formation source rock characteristics, offshore Ireland. .................................................. Appendix E.11 Figure A.E. 6 Harrier Formation source rock characteristics, offshore Ireland. ................................................. Appendix E.12 Figure A.E. 7 Inagh Formation source rock characteristics, offshore Ireland. .................................................... Appendix E.13 Figure A.E. 8 Meelagh Formation source rock characteristics, offshore Ireland. ................................................ Appendix E.14 Figure A.E. 9 Caragh Formation source rock characteristics, offshore Ireland. ................................................. Appendix E.15 Figure A.E. 10 Ruacan Formation source rock characteristics, offshore -

The Jurassic (Pliensbachian to Oxfordian) Palynology of the Algarve Basin and the Carrapateira Outlier, Southern Portugal
1 The Jurassic (Pliensbachian to Kimmeridgian) palynology of the Algarve Basin and the 2 Carrapateira outlier, southern Portugal 3 4 Marisa E.N. Borges a,b, James B. Riding c*, Paulo Fernandes a, Zélia Pereira b 5 6 a CIMA - Centro de Investigação Marinha e Ambiental. Universidade do Algarve, 7 Campus de Gambelas, 8005-139 Faro, Portugal 8 b LNEG-LGM, Rua da Amieira, 4465-965 S. Mamede Infesta, Portugal 9 c British Geological Survey, Kingsley Dunham Centre, Keyworth, Nottingham NG12 10 5GG, UK 11 12 * Corresponding author. Tel.: +44(0)115 9363447. 13 E-mail addresses: [email protected] (M.E.N. Borges), [email protected] (J.B. 14 Riding), [email protected] (P. Fernandes), [email protected] (Z. Pereira). 15 16 ABSTRACT 17 The palynology of the Jurassic (Upper Pliensbachian to Lower Kimmeridgian) 18 fill of the Algarve Basin and the Carrapateira outlier, southern Portugal was 19 investigated. Samples were collected from Armação Nova Bay, Mareta Beach, Cilheta 20 Beach and the Carrapateira outlier. At Armação Nova Bay the Upper Pliensbachian- 21 Lower Toarcian succession proved barren, or yielded sparse, non age-diagnostic 22 palynomorphs. Dinoflagellate cysts are confined to the Upper Bajocian to Upper 23 Callovian sedimentary rocks exposed at Mareta and Cilheta beaches and the Lower 24 Kimmeridgian strata of the Carrapateira outlier. At Mareta Beach, the Upper Bajocian, 25 Bathonian and Callovian produced relatively low to moderate diversity dinoflagellate 26 cyst assemblages. Several key bioevents confirm the Bathonian and Callovian ages of 27 the succession; no age-significant Late Bajocian bioevents were noted. -

Resumenes XXXI Jornadas Paleontologia.Pdf
XXXI JORNADAS DE PALEONTOLOGÍA Sociedad Española de Paleontología Baeza, 7-10 de octubre de 2015 XXXI JORNADAS DE PALEONTOLOGÍA Sociedad Española de Paleontología Baeza, 7-10 de octubre de 2015 LIBRO DE RESÚMENES Matías Reolid (ed.) Jornadas de Paleontología (31.2015.Baeza) XXXI Jornadas de Paleontología : Baeza, 7-10 de octubre de 2015 : Libro de resúmenes / [Organizan, Sociedad Española de Paleontología…[ et al.] ; Matías Reolid , (ed.) -- Jaén : Servicio de Publicaciones, Universidad de Jaén, 2015. 312 p. ; 17 x 24 cm ISBN 978-84-8439-920-9 1. Paleontología 2. Congresos y conferencias 3. Jaén (Provincia) I. Sociedad Española de Paleontología, org. II. Reolid, Matías, ed.lit . III. Universidad de Jaén. Servicio de Publicaciones, ed. IV. Título. V. Serie 566(460.352(063) XXXI JORNADAS DE PALEONTOLOGÍA Sociedad Española de Paleontología Organizan Departamento de Geología de la Universidad de Jaén Sociedad Española de Paleontología Centro de Estudios Avanzados de Ciencias de la Tierra, Universidad de Jaén Centro de Estudios de Postgrado, Universidad de Jaén Colaboran Universidad Internacional de Andalucía, Sede Antonio Machado, Baeza Diputación de Jaén Comité Español del PICG (IUGS-UNESCO) © Autores © Universidad de Jaén Primera edición, octubre 2015 Diseño y Maquetación Servicio de Publicaciones ISBN 978-84-8439-920-9 Depósito Legal J-372-2015 Edita Publicaciones de la Universidad de Jaén Vicerrectorado de Proyección de la Cultura, Deportes y Responsabilidad Social Campus Las Lagunillas, Edificio Biblioteca 23071 Jaén (España) Teléfono 953 212 355 – Fax 953 212 235 [email protected] Impreso por Gráficas “La Paz” de Torredonjimeno, S. L. Avda. de Jaén, s/n 23650 Torredonjimeno (Jaén) Teléfono 953 571 087 – Fax 953 571 207 Impreso en España / Printed in Spain “Cualquier forma de reproducción, distribución, comunicación pública o transformación de esta obra solo puede ser realizada con la autorización de sus titulares, salvo excepción prevista por la ley. -

Ne Iran, Kopet-Dagh) and Their Biostratigraphic and Palaeobiogeographic Importance
Rivista Italiana di Paleontologia e Stratigrafia (Research in Paleontology and Stratigraphy) vol. 125(2): 317-331. July 2019 SOME MICROFOSSILS (DASYCLADALES, BENTHIC FORAMINIFERA, SPONGES) FROM THE UPPER JURASSIC MOZDURAN FORMATION (NE IRAN, KOPET-DAGH) AND THEIR BIOSTRATIGRAPHIC AND PALAEOBIOGEOGRAPHIC IMPORTANCE FELIX SCHLAGINTWEIT1, ZOHREH KADIVAR2 & KOOROSH RASHIDI2 1Lerchenauerstr. 167, 80935 München, Germany. E-Mail: [email protected] 2Department of Geology, Yazd University, Safaieh, Post Box 89195-741 Yazd, Iran. E-mail: [email protected], [email protected] To cite this article: Schlagintweit F., Kadivar Z. & Rashidi K. (2019) - Some microfossils (Dasycladales, benthic foraminifera, sponges) from the Upper Jurassic Mozduran Formation (NE Iran, Kopet-Dagh) and their biostratigraphic and palaeobiogeographic importance. Riv. It. Paleontol. Strat., 125(2): 317-331. Keywords: Upper Jurassic; calcareous algae; foraminifera; sponges; shallow-water carbonates; taxonomy. Abstract. The Mozduran Formation represents mainly carbonatic shallow-water deposits from the Kopet-Dagh basin of northeast Iran. Longtime considered to be of exclusively Late Jurassic (Oxfordian-Kimmeridgian) age, its ranging into the Early Cretaceous has been demonstrated in recent times. The micropalaeontological inventory and bio- stratigraphic data however, are still poorly constrained. In the present contribution, some taxa of Dasycladales [Camp- belliella striata (Carozzi), Montenegrella florifera Bernier, Petrascula bugesiaca Bernier, Petrascula cf. bursiformis (Éttalon), Triplo- porella sp.], benthic foraminifera [Neokilianina rahonensis (Foury & Vincent), Spiraloconulus suprajurassicus Schlagintweit], and sponges (Paronadella? sp., Neuropora lusitanica G. Termier & H. Termier, Thalamopora sp.) are reported. Some taxa are reported for the first time from this formation, some even for the first time from Iran. The identified assemblage is assigned to the Tithonian, although a late Kimmeridgian age for the lowermost part of the section studied is possible. -

44. Mesozoic-Cenozoic Geology of the Eastern Margin of the Grand Banks and Its Relation to Galicia Bank1
Boillot, G., Winterer, E. L., et al., 1988 Proceedings of the Ocean Drilling Program, Scientific Results, Vol. 103 44. MESOZOIC-CENOZOIC GEOLOGY OF THE EASTERN MARGIN OF THE GRAND BANKS AND ITS RELATION TO GALICIA BANK1 A. C. Grant, L. F. Jansa, K. D. McAlpine, and A. Edwards, Geological Survey of Canada, Atlantic Geoscience Centre, Bedford Institute of Oceanography, Dartmouth, Nova Scotia, Canada ABSTRACT Late Paleozoic reconstructions of the North Atlantic juxtapose the eastern margin of the Grand Banks with the con• tinental margin off Iberia. Comparison of the geology of the Grand Banks region with results from ODP Leg 103 on the Galicia margin improves our understanding of the Mesozoic-cenozoic tectonic evolution of these regions and pro• vides new constraints on pre-drift fits. The Grand Banks region is underlain by Paleozoic and Precambrian rocks of the Appalachian Orogen, which were rifted, eroded, and buried during Mesozoic and Cenozoic tectonic episodes related to formation of the North Atlantic Ocean and the Labrador Sea. The Carson Basin along the eastern margin of the Grand Banks contains Triassic and Ju• rassic evaporites overlain by Jurassic and Cretaceous carbonate and clastic rocks that were deeply eroded during the mid-Cretaceous. This unconformity is overlain by a comparatively thin and undeformed sequence of Cretaceous-Ter• tiary fine-grained marine elastics. Since the mid-Cretaceous, the outer portion of the Carson Basin has subsided to oce• anic depths and now underlies the slope-rise zone. Comparison of the seismic stratigraphy of the Carson Basin with that of the Galicia margin indicates similar tectonic histories and depositional environments from Triassic to Tertiary time.