<I>Ariosoma Anali
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

A Review of Congrid Eels of the Genus Ariosoma from Taiwan, With
Zoological Studies 37(1): 7-12 (1998) A Review of Congrid Eels of the Genus Ariosoma from Taiwan, with Description of a New Species Shih-Chieh Shen Department of Zoology, National Taiwan University, Taipei, Taiwan 106, R.O.C. Tel: 886-2-23630231 ext. 2353. Fax: 886-2-23636837. E-mail: [email protected] (Accepted September 10, 1997) Shih-Chieh Shen (1998) A review of congrid eels of the genus Ariosoma from Taiwan, with description of a new species. Zoological Studies 37(1): 7-12. Diagnosis, description, and figures are presented for the 4 Formosan species of Ariosoma which include A. anago (Temminck and Schlegel, 1842), A. anagoides (Bleeker, 1864), A. shiroanago major (Asano, 1958) and a new species A. nancyae. Ariosoma nancyae is distinctive in its combination of a stout body; 157 total vertebrae; 53 preanal lateral-line pores, and 95 pores behind anus to tail; upper end of gill opening at level of 1/4 upper end of pectoral-fin base; black spots on head, and black bands on body, tail, and fins. Key words: Fish taxonomy, Congridae, Fish fauna, New species, Taiwan. The congrid eel genus Ariosoma Swainson, gate; preanal length more than 40% of total; tip of 1838 is not a well-known group of fishes because tail blunt and stiff, caudal fin reduced; dorsal fin of its small size and unusual habitats. It is, how- insertion origin near level of pectoral fin base; pec- ever, one of the most abundant and diverse toral fin well developed; snout rounded, projecting congrid genera in the world. -

Updated Checklist of Marine Fishes (Chordata: Craniata) from Portugal and the Proposed Extension of the Portuguese Continental Shelf
European Journal of Taxonomy 73: 1-73 ISSN 2118-9773 http://dx.doi.org/10.5852/ejt.2014.73 www.europeanjournaloftaxonomy.eu 2014 · Carneiro M. et al. This work is licensed under a Creative Commons Attribution 3.0 License. Monograph urn:lsid:zoobank.org:pub:9A5F217D-8E7B-448A-9CAB-2CCC9CC6F857 Updated checklist of marine fishes (Chordata: Craniata) from Portugal and the proposed extension of the Portuguese continental shelf Miguel CARNEIRO1,5, Rogélia MARTINS2,6, Monica LANDI*,3,7 & Filipe O. COSTA4,8 1,2 DIV-RP (Modelling and Management Fishery Resources Division), Instituto Português do Mar e da Atmosfera, Av. Brasilia 1449-006 Lisboa, Portugal. E-mail: [email protected], [email protected] 3,4 CBMA (Centre of Molecular and Environmental Biology), Department of Biology, University of Minho, Campus de Gualtar, 4710-057 Braga, Portugal. E-mail: [email protected], [email protected] * corresponding author: [email protected] 5 urn:lsid:zoobank.org:author:90A98A50-327E-4648-9DCE-75709C7A2472 6 urn:lsid:zoobank.org:author:1EB6DE00-9E91-407C-B7C4-34F31F29FD88 7 urn:lsid:zoobank.org:author:6D3AC760-77F2-4CFA-B5C7-665CB07F4CEB 8 urn:lsid:zoobank.org:author:48E53CF3-71C8-403C-BECD-10B20B3C15B4 Abstract. The study of the Portuguese marine ichthyofauna has a long historical tradition, rooted back in the 18th Century. Here we present an annotated checklist of the marine fishes from Portuguese waters, including the area encompassed by the proposed extension of the Portuguese continental shelf and the Economic Exclusive Zone (EEZ). The list is based on historical literature records and taxon occurrence data obtained from natural history collections, together with new revisions and occurrences. -

Redalyc.Congridae Congrios
Revista de Biología Tropical ISSN: 0034-7744 [email protected] Universidad de Costa Rica Costa Rica Congridae congrios Revista de Biología Tropical, vol. 58, núm. 2, octubre, 2010, pp. 30-31 Universidad de Costa Rica San Pedro de Montes de Oca, Costa Rica Disponible en: http://www.redalyc.org/articulo.oa?id=44921017036 Cómo citar el artículo Número completo Sistema de Información Científica Más información del artículo Red de Revistas Científicas de América Latina, el Caribe, España y Portugal Página de la revista en redalyc.org Proyecto académico sin fines de lucro, desarrollado bajo la iniciativa de acceso abierto HETERENCHELYIDAE HETERENCHELYIDAE anguilas de fango mud eels Pythonichthys sanguineus, anguila del fango – Ojo diminuto; Pythonichthys sanguineus, atlantic mud eel – Eye minute; mandíbula inferior proyectándose hacia adelante de la supe- lower jaw projecting beyond upper; pectoral fins and lateral rior; aletas pectorales y línea lateral ausentes; aletas dorsal y line absent; dorsal and anal fins low and confluent with caudal anal bajas y confluentes con la aleta caudal (UCR 220-26). fin. MURAENESOCIDAE MURAENESOCIDAE congrios picudos pike congers Cynoponticus savanna, congrio espantoso – Morro se proyec- Cynoponticus savanna, conehead eel – Snout projects beyond ta hacia delante de la mandíbula inferior; aberturas branquiales lower jaw; large gill openings nearly meet ventrally; lateral grandes casi se juntan ventralmente; línea lateral completa; line complete; dorsal fin origin above pectoral fin; pectoral fin origen -

Diet of a Mediterranean Monk Seal Monachus Monachus in a Transitional Post-Weaning Phase and Its Implications for the Conservation of the Species
Vol. 39: 315–320, 2019 ENDANGERED SPECIES RESEARCH Published August 22 https://doi.org/10.3354/esr00971 Endang Species Res OPENPEN ACCESSCCESS NOTE Diet of a Mediterranean monk seal Monachus monachus in a transitional post-weaning phase and its implications for the conservation of the species Cem Orkun Kıraç1,*, Meltem Ok2 1Underwater Research Society - Mediterranean Seal Research Group (SAD-AFAG), 06570 Ankara, Turkey 2Middle East Technical University - Institute of Marine Science (METU-IMS), Erdemli, 33731 Mersin, Turkey ABSTRACT: The Mediterranean monk seal Monachus monachus is the most endangered pin- niped in the world and is considered Endangered by the IUCN. Transition from suckling to active feeding is a critical time in the development of all mammal species, and understanding the dietary requirements of seals during this vulnerable period is of value in establishing conservation meas- ures, such as fishery regulations. This study provides unique information on the dietary habits of a moulted monk seal pup, through the opportunistic necropsy of a dead animal encountered at a very early age (5 mo). A total of 6 prey items from 2 families (Octopodidae, 90.8% and Congridae, 8.9%) were identified from stomach contents. The remaining stomach content mass consisted of fish bones from unidentified species (0.3%). The estimated age, low diversity and number of prey items in the stomach contents indicate that this individual may have been in a transition period from suckling to active feeding. The study confirms independent foraging in Mediterranean monk seals at about 5 mo of age. Given the importance of early life survival for maintaining stable Medi- terranean monk seal populations, and the occurrence of an ontogenetic shift in its close relative (Hawaiian monk seal), these findings contribute to the establishment and implementation of suc- cessful conservation and management strategies for this Endangered species. -

Annotated Checklist of the Fishes of Lord Howe Island
AUSTRALIAN MUSEUM SCIENTIFIC PUBLICATIONS Allen, Gerald R., Douglass F. Hoese, John R. Paxton, J. E. Randall, C. Russell, W. A. Starck, F. H. Talbot, and G. P. Whitley, 1977. Annotated checklist of the fishes of Lord Howe Island. Records of the Australian Museum 30(15): 365–454. [21 December 1976]. doi:10.3853/j.0067-1975.30.1977.287 ISSN 0067-1975 Published by the Australian Museum, Sydney naturenature cultureculture discover discover AustralianAustralian Museum Museum science science is is freely freely accessible accessible online online at at www.australianmuseum.net.au/publications/www.australianmuseum.net.au/publications/ 66 CollegeCollege Street,Street, SydneySydney NSWNSW 2010,2010, AustraliaAustralia ANNOTATED CHECKLIST OF THE FISHES OF LORD HOWE ISLAND G. R. ALLEN, 1,2 D. F. HOESE,1 J. R. PAXTON,1 J. E. RANDALL, 3 B. C. RUSSELL},4 W. A. STARCK 11,1 F. H. TALBOT,1,4 AND G. P. WHITlEy5 SUMMARY lord Howe Island, some 630 kilometres off the northern coast of New South Wales, Australia at 31.5° South latitude, is the world's southern most locality with a well developed coral reef community and associated lagoon. An extensive collection of fishes from lord Howelsland was made during a month's expedition in February 1973. A total of 208 species are newly recorded from lord Howe Island and 23 species newly recorded from the Australian mainland. The fish fauna of lord Howe is increased to 447 species in 107 families. Of the 390 species of inshore fishes, the majority (60%) are wide-ranging tropical forms; some 10% are found only at lord Howe Island, southern Australia and/or New Zealand. -

Anguilliformes, Saccopharyngiformes, and Notacanthiformes (Teleostei: Elopomorpha)
* Catalog of Type Specimens of Recent Fishes in the National Museum of Natural History, Smithsonian Institution, 6: Anguilliformes, Saccopharyngiformes, and Notacanthiformes (Teleostei: Elopomorpha) DAVID G. SMITH I SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY • NUMBER 566 SERIES PUBLICATIONS OF THE SMITHSONIAN INSTITUTION Emphasis upon publication as a means of "diffusing knowledge" was expressed by the first Secretary of the Smithsonian. In his formal plan for the Institution, Joseph Henry outlined a program that included the following statement: "It is proposed to publish a series of reports, giving an account of the new discoveries in science, and of the changes made from year to year in all branches of knowledge." This theme of basic research has been adhered to through the years by thousands of titles issued in series publications under the Smithsonian imprint, commencing with Smithsonian Contributions to Knowledge in 1848 and continuing with the following active series: Smithsonian Contributions to Anthropology Smithsonian Contributions to Astrophysics Smithsonian Contributions to Botany Smithsonian Contributions to the Earth Sciences Smithsonian Contributions to the Marine Sciences Smithsonian Contributions to Paleobiology Smithsonian Contributions to Zoology Smithsonian Folklife Studies Smithsonian Studies in Air and Space Smithsonian Studies in History and Technology In these series, the Institution publishes small papers and full-scale monographs that report the research and collections of its various museums and bureaux or of professional colleagues in the world of science and scholarship. The publications are distributed by mailing lists to libraries, universities, and similar institutions throughout the world. Papers or monographs'submitted for series publication are received by the Smithsonian Institution Press, subject to its own review for format and style, only through departments of the various Smithsonian museums or bureaux, where the manuscripts are given substantive review. -
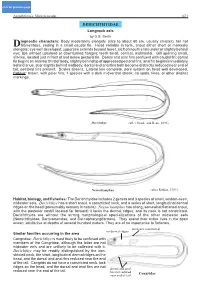
DERICHTHYIDAE Longneck Eels by D.G
click for previous page Anguilliformes: Muraenesocidae 1671 DERICHTHYIDAE Longneck eels by D.G. Smith iagnostic characters: Body moderately elongate (size to about 60 cm, usually smaller), tail not Dfilamentous, ending in a small caudal fin. Head variable in form, snout either short or markedly elongate; eye well developed; upper jaw extends beyond lower, cleft of mouth ends under or slightly behind eye; lips without upturned or downturned flanges; teeth small, conical, multiserial. Gill opening small, slit-like, located just in front of and below pectoral fin. Dorsal and anal fins confluent with caudal fin; dorsal fin begins on anterior third of body, slightly behind tip of appressed pectoral fins; anal fin begins immediately behind anus, at or slightly behind midbody; dorsal and anal fins both become distinctly reduced near end of tail; pectoral fins present. Scales absent. Lateral line complete, pore system on head well developed. Colour: brown, with paler fins; 1 species with a dark midventral streak; no spots, lines, or other distinct markings. Derichthys (after Goode and Bean, 1896) Nessorhamphus (after Robins, 1989) Habitat, biology, and fisheries: The Derichthyidae includes 2 genera and 3 species of small, seldom-seen, midwater eels. Derichthys has a short snout, a constricted neck, and a series of short, longitudinal dermal ridges on the head (presumably sensory in nature). Nessorhamphus has a long, somewhat flattened snout, with the posterior nostril located far forward; it lacks the dermal ridges, and its neck is not constricted. Derichthyids are without the strong morphological specializations of the other midwater eels (Nemichthyidae, Serrivomeridae, and Saccopharyngiformes). They spend their entire lives in the open ocean; adults live at depths of several hundred meters. -

Elopomorph Larvae Are Important Contributors to Fish Biodiversity in a Low-Latitude Oceanic Ecosystem
fmars-07-00169 April 27, 2020 Time: 22:6 # 1 ORIGINAL RESEARCH published: 29 April 2020 doi: 10.3389/fmars.2020.00169 Hiding in Plain Sight: Elopomorph Larvae Are Important Contributors to Fish Biodiversity in a Low-Latitude Oceanic Ecosystem Jon A. Moore1,2*, Dante B. Fenolio3, April B. Cook4 and Tracey T. Sutton4 1 Harriet L. Wilkes Honors College, Florida Atlantic University, Jupiter, FL, United States, 2 Harbor Branch Oceanographic Institute, Florida Atlantic University, Fort Pierce, FL, United States, 3 Center for Conservation and Research, San Antonio Zoo, San Antonio, TX, United States, 4 Halmos College of Natural Sciences and Oceanography, Nova Southeastern University, Dania Beach, FL, United States Leptocephalus larvae of elopomorph fishes are a cryptic component of fish diversity in nearshore and oceanic habitats. However, identifying those leptocephali can be important in illuminating species richness in a region. Since the Deepwater Horizon oil spill in 2010, sampling of offshore fishes in the epi-, meso-, and upper bathypelagic Edited by: depth strata of the northern Gulf of Mexico resulted in 8989 identifiable specimens of Michael Vecchione, leptocephalus larvae or transforming juveniles, in 118 taxa representing 83 recognized National Oceanic and Atmospheric Administration (NOAA), United States and established species and an additional 35 distinctive leptocephalus morphotypes Reviewed by: not yet linked to a known described species. Leptocephali account for ∼13% of the Mackenzie E. Gerringer, total species richness of fishes collected in the offshore region. A new morphotype SUNY Geneseo, United States Dave Johnson, of Muraenidae leptocephalus is also described. We compare this study with other National Museum of Natural History leptocephalus diversity studies in the western Atlantic. -

Species Acromycter Nezumi (Asano, 1958)
FAMILY Congridae Kaup, 1856 - conger eels SUBFAMILY Congrinae Kaup, 1856 - conger eels [=Atteridi, Ophioides, Leptocephalini, Congeridae, Macrocephenchelyidae, Scalanogoinae, Urocongrinae] GENUS Acromycter Smith & Kanazawa, 1977 - conger eels Species Acromycter alcocki (Gilbert & Cramer, 1897) - Alcock's congrid eel Species Acromycter atlanticus Smith, 1989 - Atlantic conger eel Species Acromycter longipectoralis Karmovskaya, 2004 - longfinned conger eel Species Acromycter nezumi (Asano, 1958) - Nezumi's conger eel Species Acromycter perturbator (Parr, 1932) - perturbator conger eel GENUS Bassanago Whitley, 1948 - conger eels Species Bassanago albescens (Barnard, 1923) - hairy conger Species Bassanago bulbiceps Whitley, 1948 - swollen-headed conger eel Species Bassanago hirsutus (Castle, 1960) - deepsea conger Species Bassanago nielseni (Karmovskaya, 1990) - Nielsen's conger eel GENUS Bathycongrus Ogilby, 1898 - conger eels [=Congrina, Microcephalocongrus, Pseudoxenomystax, Rhechias, Uranoconger] Species Bathycongrus aequoreus (Gilbert & Cramer, 1897) - Kaiwi bathycongrus [=armiger] Species Bathycongrus bertini (Poll, 1953) - Poll's conger Species Bathycongrus bleekeri Fowler, 1934 - Bleeker's bathycongrus Species Bathycongrus bullisi (Smith & Kanazawa, 1977) - bullish conger Species Bathycongrus dubius (Breder, 1927) - dubius conger Species Bathycongrus guttulatus (Günther, 1887) - lined conger [=roosendaali, stimpsoni] Species Bathycongrus longicavis Karmovskaya, 2009 - Vanuatu conger Species Bathycongrus macrocercus (Alcock, 1894) - -

The Sargasso Sea Subtropical Gyre the Spawning and Larval Development Area of Both Freshwater and Marine Eels
The Sargasso Sea Subtropical Gyre The Spawning and Larval Development Area of Both Freshwater and Marine Eels Michael J. Miller and Reinhold Hanel Number 7 Sargasso Sea Alliance Science Report Series When referenced this report should be referred to as: Miller, M.J. and R. Hanel. 2011. The Sargasso Sea Subtropical Gyre: The Spawning and Larval Development Area of both Freshwater and Marine eels. Sargasso Sea Alliance Science Report Series, No 7, 20 pp. ISBN 978-0-9847520-9-6. The Sargasso Sea Alliance is led by the Bermuda Government and aims to promote international awareness of the importance of the Sargasso Sea and to mobilise support from a wide variety of national and international organisations, governments, donors and users for protection measures for the Sargasso Sea. Further details: Dr David Freestone, Executive Director, Sargasso Sea Alliance, Suite 300, 1630 Connecticut Avenue NW, Washington D.C., 20009, USA. Email: [email protected] Kate K. Morrison, Deputy Director, at the same address Email: [email protected] The Secretariat of the Sargasso Sea Alliance is hosted by the Washington D.C. Office of the International Union for the Conservation of Nature (IUCN). Website is www.sargassoalliance.org This case is being produced with generous support of donors to the Sargasso Sea Alliance: Ricardo Cisneros, Erik H. Gordon, JM Kaplan Fund, Richard Rockefeller, David E. Shaw, and the Waitt Foundation. Additional support provided by: WWF Sweden and the Pew Environment Group. COVER photo: European eel, Reinhold Hanel. ISBN 978-0-9847520-9-6 The Sargasso Sea Subtropical Gyre The Spawning and Larval Development Area of Both Freshwater and Marine Eels Michael J. -
Status of Non-CITES Listed Anguillid Eels
Status of non-CITES listed anguillid eels Matthew Gollock, Hiromi Shiraishi, Savrina Carrizo, Vicki Crook and Emma Levy Delivered by the Zoological Society of London 1 Acknowledgments This report was made possible with financial support from the CITES Secretariat. The authors would like to thank Karen Gaynor in the CITES Secretariat for support during the drafting of the report. We thank from TRAFFIC, Joyce Wu for supporting the study and James Compton and Richard Thomas for reviewing elements of the draft report. Camilla Beevor and Aaron Foy at ZSL are thanked for their legal and operational support, Kristen Steele for translation of documents, Joanna Barker for creating the range maps and David Curnick for editing a draft of the report. Kenzo Kaifu and Céline Hanzen are acknowledged for technical input. Parties are thanked for completing the questionnaires that informed this report and for reviewing a draft to ensure it is as accurate and up to date as possible. We also thank the individuals who responded to specific requests for information and clarification. Author affiliations Matthew Gollock and Emma Levy, Zoological Society of London, Regent’s Park, London, NW1 4RY, United Kingdom. Hiromi Shiraishi, TRAFFIC, The David Attenborough Building, Pembroke Street, Cambridge CB2 3QZ, United Kingdom. Savrina Carrizo - Independent consultant. Vicki Crook – Independent consultant. 2 Table of Contents Executive summary ............................................................................................................................... -

Larval Size-Distributions of Ariosoma Balearicum Cryptic Species During the March–April Season in the Sargasso Sea Subtropical Convergence Zone
1 Environmental Biology of Fishes Archimer October 2019, Volume 102, Issue 10, Pages 1231-1252 https://doi.org/10.1007/s10641-019-00900-8 https://archimer.ifremer.fr https://archimer.ifremer.fr/doc/00509/62028/ Larval size-distributions of Ariosoma balearicum cryptic species during the March–April season in the Sargasso Sea Subtropical Convergence Zone Miller Michael J. 1, 2, * , Marohn Lasse 3, Wysujack Klaus 3, Bonhommeau Sylvain 4, Kuroki Mari 2, Freese Marko 3, Pohlmann Jan-Dag 3, Watanabe Shun 5, Blancke Tina 3, Weist Peggy 3, Castonguay Martin 6, Westerberg Håkan 7, Tsukamoto Katsumi 1, 2, Hanel Reinhold 3 1 Department of Marine Science and Resources, College of Bioresource SciencesNihon UniversityFujisawa, Japan 2 Department of Aquatic Bioscience, Graduate School of Agricultural and Life SciencesThe University of TokyoTokyo, Japan 3 Thünen Institute of Fisheries EcologyBremerhaven, Germany 4 IfremerLe Port Cedex, France 5 Department of Fisheries, Faculty of AgricultureKindai UniversityNara, Japan 6 Fisheries and Oceans CanadaInstitut Maurice-LamontagneMont-Joli, Canada 7 Department of Aquatic Resources, Institute of Freshwater ResearchSwedish University of Agricultural SciencesDrottningholm, Sweden * Corresponding author : Michael J. Miller, email address : [email protected] Abstract : Leptocephali of the shallow-water congrid eel Ariosoma balearicum are abundant during February–April in the Sargasso Sea, and larval and adult meristic data indicates this species includes several regional subpopulations/cryptic species. Four multiple-transect larval surveys (2011, 2014, 2015, 2017) were used to examine the geographic size distribution of two myomere-count types of A. balearicum leptocephali. High-count (HC) larvae were consistently mostly between 80 and 100 mm in size (60–132 mm; 87.9 ± 6.8 mm) as observed previously, and frequently had narrow size ranges.