Aftermath of a Feast : Human Colonization of the Southern Bahamian Archipelago and Its Effects on the Indigenous Fauna
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Iguanas and Seabirds
PROTECTEDAREASMANAGEMENTSTRATEGYFOR BAHAMIAN TERRESTRIAL VERTEBRATES: IGUANAS AND SEABIRDS Bahamian Field Station San Salvador, The Bahamas 11-12 November, 2000 Organized by Conservation Unit, Bahamas Department of Agriculture and IUCN/SSC Iguana Specialist Group In collaboration with IUCN/SSC Conservation Breeding Specialist Group Supported by Fort Worth Zoo Zoological Society of San Diego A contribution of the IUCN/SSC Conservation Breeding Specialist Group. Organized by Conservation Unit, Bahamas Department of Agriculture and the IUCN/SSC Iguana Specialist Group, in collaboration with the IUCN/SSC Conservation Breeding Specialist Group. Supported by the Fort Worth Zoo and the Zoological Society of San Diego. © Copyright 2001 by CBSG. Citation: E. Carey, S.D. Buckner, A. C. Alberts, R.D. Hudson, and D. Lee, editors. 2001. Protected Areas Management Strategy for Bahamian Terrestrial Vertebrates: Iguanas and Seabirds. IUCN/SSC Conservation Breeding Specialist Group, Apple Valley, Minnesota. Additional copies of Protected Areas Management Strategy for Bahamian Terrestrial Vertebrates: Iguanas and Seabirds Report can be ordered through the the IUCN/SSC Conservation Breeding Specialist Group, 12101 Johnny Cake Ridge Road, Apple Valley, MN 55124. Bartschi s iguana, Cyclura carinata bartschi Andros island iguana, Cyclura cychlura cychlura Exuma island iguana, Cyclura cychlura figginsi Allen s Cay iguana, Cyclura cychlura inornata Allen s Cay iguana, Cyclura cychlura inornata Acklins iguana, Cyclura rileyi nucha/is San Salvador iguana, Cyclura rileyi rileyi San Salvador iguana, Cyclura rileyi rileyi Audubon s Shearwater, Puffinus lherminieri Least Tern, Sterna antillarum White-tailed Tropicbird, Phaethon lepturus Brown Booby, Sula leucogaster Bridled Tern, Sterna anaethetus Magnificent Frigatebird, Fregata magnificens - Juveniles CONTENTS Opening Remarks by The Bahamas Minister of Commerce, Agriculture, and Industry ......................... -

WMSDB - Worldwide Mollusc Species Data Base
WMSDB - Worldwide Mollusc Species Data Base Family: TURBINIDAE Author: Claudio Galli - [email protected] (updated 07/set/2015) Class: GASTROPODA --- Clade: VETIGASTROPODA-TROCHOIDEA ------ Family: TURBINIDAE Rafinesque, 1815 (Sea) - Alphabetic order - when first name is in bold the species has images Taxa=681, Genus=26, Subgenus=17, Species=203, Subspecies=23, Synonyms=411, Images=168 abyssorum , Bolma henica abyssorum M.M. Schepman, 1908 aculeata , Guildfordia aculeata S. Kosuge, 1979 aculeatus , Turbo aculeatus T. Allan, 1818 - syn of: Epitonium muricatum (A. Risso, 1826) acutangulus, Turbo acutangulus C. Linnaeus, 1758 acutus , Turbo acutus E. Donovan, 1804 - syn of: Turbonilla acuta (E. Donovan, 1804) aegyptius , Turbo aegyptius J.F. Gmelin, 1791 - syn of: Rubritrochus declivis (P. Forsskål in C. Niebuhr, 1775) aereus , Turbo aereus J. Adams, 1797 - syn of: Rissoa parva (E.M. Da Costa, 1778) aethiops , Turbo aethiops J.F. Gmelin, 1791 - syn of: Diloma aethiops (J.F. Gmelin, 1791) agonistes , Turbo agonistes W.H. Dall & W.H. Ochsner, 1928 - syn of: Turbo scitulus (W.H. Dall, 1919) albidus , Turbo albidus F. Kanmacher, 1798 - syn of: Graphis albida (F. Kanmacher, 1798) albocinctus , Turbo albocinctus J.H.F. Link, 1807 - syn of: Littorina saxatilis (A.G. Olivi, 1792) albofasciatus , Turbo albofasciatus L. Bozzetti, 1994 albofasciatus , Marmarostoma albofasciatus L. Bozzetti, 1994 - syn of: Turbo albofasciatus L. Bozzetti, 1994 albulus , Turbo albulus O. Fabricius, 1780 - syn of: Menestho albula (O. Fabricius, 1780) albus , Turbo albus J. Adams, 1797 - syn of: Rissoa parva (E.M. Da Costa, 1778) albus, Turbo albus T. Pennant, 1777 amabilis , Turbo amabilis H. Ozaki, 1954 - syn of: Bolma guttata (A. Adams, 1863) americanum , Lithopoma americanum (J.F. -

ABSTRACT Title of Dissertation: PATTERNS IN
ABSTRACT Title of Dissertation: PATTERNS IN DIVERSITY AND DISTRIBUTION OF BENTHIC MOLLUSCS ALONG A DEPTH GRADIENT IN THE BAHAMAS Michael Joseph Dowgiallo, Doctor of Philosophy, 2004 Dissertation directed by: Professor Marjorie L. Reaka-Kudla Department of Biology, UMCP Species richness and abundance of benthic bivalve and gastropod molluscs was determined over a depth gradient of 5 - 244 m at Lee Stocking Island, Bahamas by deploying replicate benthic collectors at five sites at 5 m, 14 m, 46 m, 153 m, and 244 m for six months beginning in December 1993. A total of 773 individual molluscs comprising at least 72 taxa were retrieved from the collectors. Analysis of the molluscan fauna that colonized the collectors showed overwhelmingly higher abundance and diversity at the 5 m, 14 m, and 46 m sites as compared to the deeper sites at 153 m and 244 m. Irradiance, temperature, and habitat heterogeneity all declined with depth, coincident with declines in the abundance and diversity of the molluscs. Herbivorous modes of feeding predominated (52%) and carnivorous modes of feeding were common (44%) over the range of depths studied at Lee Stocking Island, but mode of feeding did not change significantly over depth. One bivalve and one gastropod species showed a significant decline in body size with increasing depth. Analysis of data for 960 species of gastropod molluscs from the Western Atlantic Gastropod Database of the Academy of Natural Sciences (ANS) that have ranges including the Bahamas showed a positive correlation between body size of species of gastropods and their geographic ranges. There was also a positive correlation between depth range and the size of the geographic range. -

The Last Survivors: Current Status and Conservation of the Non-Volant Land
1 The Last Survivors: current status and conservation of the non-volant land 2 mammals of the insular Caribbean 3 4 SAMUEL T. TURVEY,* ROSALIND J. KENNERLEY, JOSE M. NUÑEZ-MIÑO, AND RICHARD P. 5 YOUNG 6 7 Institute of Zoology, Zoological Society of London, Regent’s Park, London NW1 4RY (STT) 8 Durrell Wildlife Conservation Trust, Les Augrès Manor, Trinity, Jersey JE3 5BP, Channel 9 Islands (RJK, JNM, RPY) 10 11 *Correspondent: [email protected] 12 13 Running header: Status of Caribbean land mammals 14 1 15 The insular Caribbean is among the few oceanic-type island systems colonized by non-volant 16 land mammals. This region also has experienced the world’s highest levels of historical 17 mammal extinctions, with at least 29 species lost since AD 1500. Representatives of only 2 18 land-mammal families (Capromyidae and Solenodontidae) now survive, in Cuba, Hispaniola, 19 Jamaica, and the Bahama Archipelago. The conservation status of Caribbean land mammals 20 is surprisingly poorly understood. The most recent IUCN Red List assessment, from 2008, 21 recognized 15 endemic species, of which 13 were assessed as threatened. We reassessed all 22 available baseline data on the current status of the Caribbean land-mammal fauna within the 23 framework of the IUCN Red List, to determine specific conservation requirements for 24 Caribbean land-mammal species using an evidence-based approach. We recognize only 13 25 surviving species, 1 of which is not formally described and cannot be assessed using IUCN 26 criteria; 3 further species previously considered valid are interpreted as junior synonyms or 27 subspecies. -

Marine Shells of the Western Coast of Flordia
wm :iii! mm ilili ! Sfixing cHdL J^oad .Sandivicl'i, j\{ai.i.ach.u±£.tti. icuxucm \^*^£ FRONTISPIECE Photo by Ruth Bernhard Spondylus americanus Hermann MARINE SHELLS f>4 OF THE WESTERN COAST OF FLORIDA By LOUISE M. PERRY AND JEANNE S. SCHWENGEL With Revisions and Additions to Louise M. Perry's Marine Shells of the Southwest Coast of Florida Illustrations by W. Hammersley Southwick, Axel A. Olsson, and Frank White March, 1955 PALEONTOLOGICAL RESEARCH INSTITUTION ITHACA, NEW YORK U. S. A. MARINE SHELLS OF THE SOUTHWEST COAST OF FLORIDA printed as Bulletins of American Paleontology, vol. 26, No. 95 First printing, 1940 Second printing, 1942 Copyright, 1955, by Paleontological Research Institution Library of Congress Catalog Card Number: 5-^-12005 Printed in the United States of America // is perhaps a more fortunate destiny to have a taste for collecting shells than to be born a millionaire. Robert Louis Stevenson imeters 50 lllllllllllllllllllllllllllll II II III nil 2 Inches CONTENTS Page Preface by reviser 7 Foreword by Wm. J. Clench 9 Introduction 11 Generalia 13 Collection and preparation of specimens 17 Systematic descriptions 24 Class Amphineura :. 24 Class Pelecypoda 27 Class Scaphopoda 97 Class Gasteropoda 101 Plates 199 Index 311 PREFACE BY THE REVISER It has been a privilege to revise Louise M. Perry's fine book on "Marine Shells of Southwest Florida", to include her studies on eggs and larvae of mollusks; and to add descriptions and illustra- tions of several newly discovered shells thus making it a more com- prehensive study of the molluscan life of western Florida. The work that I have done is only a small return to Dr. -

Mollusks of Candomblé: Symbolic and Ritualistic Importance Nivaldo a Léo Neto1,3, Robert a Voeks2, Thelma LP Dias3 and Rômulo RN Alves3*
Léo Neto et al. Journal of Ethnobiology and Ethnomedicine 2012, 8:10 http://www.ethnobiomed.com/content/8/1/10 JOURNAL OF ETHNOBIOLOGY AND ETHNOMEDICINE RESEARCH Open Access Mollusks of Candomblé: symbolic and ritualistic importance Nivaldo A Léo Neto1,3, Robert A Voeks2, Thelma LP Dias3 and Rômulo RN Alves3* Abstract Human societies utilize mollusks for myriad material and spiritual ends. An example of their use in a religious context is found in Brazil’s African-derived belief systems. Candomblé, an Afro-Brazilian religion introduced during the 18th-19th centuries by enslaved Yoruba, includes various magical and liturgical uses of mollusks. This work inventoried the species utilized by adherents and to analyzed their symbolic and magical context. Data were obtained from Candomblé temples in two cities in the northeast of Brazil-Caruaru, in the state of Pernambuco, and Campina Grande, in the state of Paraíba. Questionnaires administered to eleven adepts revealed that at least nineteen mollusk species are being used. Shells from Monetaria moneta, M. annulus and Erosaria caputserpentis were cited by all of the interviewees. Three uses stood out: divination (jogo de búzios); utilization as ritual objects; and employment as sacrificial offerings (Igbin or Boi-de-Oxalá). The jogo de búzios (shell toss), employed in West Africa, Brazil and Cuba, is of fundamental importance to the cult, representing the means by which the faithful enter in contact with the divinities (Orixás) and consult people’s futures (Odu). The utilization of mollusks in Candomblé is strongly influenced by ancient Yoruba myths (Itãs) which, having survived enslavement and generations of captive labor, continue to guide the lives of Brazil’s African Diaspora. -

Florida Keys Species List
FKNMS Species List A B C D E F G H I J K L M N O P Q R S T 1 Marine and Terrestrial Species of the Florida Keys 2 Phylum Subphylum Class Subclass Order Suborder Infraorder Superfamily Family Scientific Name Common Name Notes 3 1 Porifera (Sponges) Demospongia Dictyoceratida Spongiidae Euryspongia rosea species from G.P. Schmahl, BNP survey 4 2 Fasciospongia cerebriformis species from G.P. Schmahl, BNP survey 5 3 Hippospongia gossypina Velvet sponge 6 4 Hippospongia lachne Sheepswool sponge 7 5 Oligoceras violacea Tortugas survey, Wheaton list 8 6 Spongia barbara Yellow sponge 9 7 Spongia graminea Glove sponge 10 8 Spongia obscura Grass sponge 11 9 Spongia sterea Wire sponge 12 10 Irciniidae Ircinia campana Vase sponge 13 11 Ircinia felix Stinker sponge 14 12 Ircinia cf. Ramosa species from G.P. Schmahl, BNP survey 15 13 Ircinia strobilina Black-ball sponge 16 14 Smenospongia aurea species from G.P. Schmahl, BNP survey, Tortugas survey, Wheaton list 17 15 Thorecta horridus recorded from Keys by Wiedenmayer 18 16 Dendroceratida Dysideidae Dysidea etheria species from G.P. Schmahl, BNP survey; Tortugas survey, Wheaton list 19 17 Dysidea fragilis species from G.P. Schmahl, BNP survey; Tortugas survey, Wheaton list 20 18 Dysidea janiae species from G.P. Schmahl, BNP survey; Tortugas survey, Wheaton list 21 19 Dysidea variabilis species from G.P. Schmahl, BNP survey 22 20 Verongida Druinellidae Pseudoceratina crassa Branching tube sponge 23 21 Aplysinidae Aplysina archeri species from G.P. Schmahl, BNP survey 24 22 Aplysina cauliformis Row pore rope sponge 25 23 Aplysina fistularis Yellow tube sponge 26 24 Aplysina lacunosa 27 25 Verongula rigida Pitted sponge 28 26 Darwinellidae Aplysilla sulfurea species from G.P. -

Caenogastropoda: Hydrobiidae) in the Caucasus
Folia Malacol. 25(4): 237–247 https://doi.org/10.12657/folmal.025.025 AGRAFIA SZAROWSKA ET FALNIOWSKI, 2011 (CAENOGASTROPODA: HYDROBIIDAE) IN THE CAUCASUS JOZEF GREGO1, SEBASTIAN HOFMAN2, LEVAN MUMLADZE3, ANDRZEJ FALNIOWSKI4* 1Horná Mičiná 219, 97401 Banská Bystrica, Slovakia 2Department of Comparative Anatomy, Institute of Zoology, Jagiellonian University, Cracow, Poland 3Institute of Zoology Ilia State University, Tbilisi, Georgia 4Department of Malacology, Institute of Zoology, Jagiellonian University, Cracow, Poland (e-mail: [email protected]) *corresponding author ABSTRACT: Freshwater gastropods of the Caucasus are poorly known. A few minute Belgrandiella-like gastropods were found in three springs in Georgia. Molecular markers: mitochondrial cytochrome oxidase subunit I (COI) and nuclear histone (H3) were used to infer their phylogenetic relationships. The phylogenetic trees placed them most closely to Agrafia from continental Greece. The p-distances indicated that two species occurred in the three localities. Two specimens from Andros Island (Greece) were also assigned to the genus Agrafia. The p-distances between the four taxa, most probably each representing a distinct species, were within the range of 0.026–0.043 for H3, and 0.089–0.118 for COI. KEY WORDS: spring snail, DNA, Georgia, Andros, phylogeny INTRODUCTION The freshwater gastropods of Georgia, as well as Graziana Radoman, 1975, Pontobelgrandiella Radoman, those of all the Caucasus, are still poorly studied; this 1973, and Alzoniella Giusti et Bodon, 1984. They are concerns especially the minute representatives of the morphologically distinguishable but only slightly Truncatelloidea. About ten species inhabiting karst different, and molecularly represent not closely re- springs and caves have been found so far (SHADIN lated lineages. -

A Survey of the Littoral Gastropoda of the Netherlands Antilles And
STUDIES ON THE FAUNA OF CURAÇAO AND OTHER CARIBBEAN ISLANDS: No. 31 A Survey of the Littoral Gastropoda of the Netherlands Antilles and other Caribbean Islands by H.E. Coomans (Zoologisch Museum, Amsterdam) This brief is based the material Dr. survey on collected by P. WagenaarHummelinck in 1936/37, 1948/49, and 1955. Station numbers only are cited; they refer to the “Description of new localities” in Volume IV of this series (1953; marine habitats p. 56-77) and to a “Third list oflocalities”which willbe published in a forthcoming volume. Other localities, which are not numbered, described in the are, as a rule, briefly text. Material assembled by a few other collectors has been added to Hummelinck’s collection. The names of the collectors are always mentioned, abbreviated as follows: Av.: R. Aveledo, Caracas Ga.: Wiesje and Hendrikje, Be.: J. G. van den Bergh, Aruba the two little daughters of BL. : T. Blok, Curaçao Mr. and Mrs. K. J. van Bo.: Mrs van den Bos, St Maarten Gaalen, Aruba. Co.: R. M. Collens, Tobago Za. : J. S. Zaneveld, Curaçao. For the sake of completeness the material discussed by TERA VAN BENTHEM in her "Marine Molluscs of the JUTTING paper on Island of Curasao" (1927) has been included (indicated by V.B.J.), few together with a other data on Netherlands Antilles Gastropoda from the literature of the subject, including those mentioned by J. J. SCHEPMAN in his contribution (1915) to the Encyclopaedie van Nederlandsch West-Indie (indicated by SCH.). 43 of his Dr. HUMMELINCK informed me that a considerable part marine material has not yet been examined for the presence of Gastropoda, and that the majority of the samples collected by Dr. -
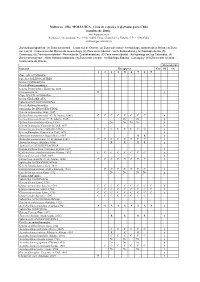
Moluscos - Filo MOLLUSCA
Moluscos - Filo MOLLUSCA. Lista de especies registradas para Cuba (octubre de 2006). José Espinosa Sáez Instituto de Oceanología, Ave 1ª No. 18406, Playa, Ciudad de La Habana, C.P. 11200, Cuba [email protected] Zonas biogeográficas: (1) Zona suroriental – Costa sur de Oriente, (2) Zona surcentral - Archipiélago Jardines de la Reina, (3) Zona sur central - Costa al sur del Macizo de Guamuhaya, (4) Zona suroccidental - Golfo de Batabanó y Archipiélago de los, (5) Canarreos, (6) Zona suroccidental - Península de Guanahacabibes, (7) Zona noroccidental - Archipiélago de Los Colorados, (8) Zona noroccidental - Norte Habana-Matanzas, (9) Zona norte-central - Archipiélago Sabana - Camagüey, (10) Zona norte-oriental - Costa norte de Oriente Abreviaturas Especies Bioegiones Cu Pl Oc 1 2 3 4 5 6 7 8 9 Clase APLACOPHORA Subclase SOLENOGASTRES Orden CAVIBELONIA Familia Proneomeniidae Género Proneomenia Hubrecht, 1880 Proneomenia sp . R x Clase POLYPLACOPHORA Orden NEOLORICATA Suborden ISCHNOCHITONINA Familia Ischnochitonidae Subfamilia ISCHNOCHITONINAE Género Ischnochiton Gray, 1847 Ischnochiton erythronotus (C. B. Adams, 1845) C C C C C C C C x Ischnochiton papillosus (C. B. Adams, 1845) Nc Nc x Ischnochiton striolatus (Gray, 1828) Nc Nc Nc Nc x Género Ischnoplax Carpenter in Dall, 1879 x Ischnoplax pectinatus (Sowerby, 1832) C C C C C C C C x Género Stenoplax Carpenter in Dall, 1879 x Stenoplax bahamensis Kaas y Belle, 1987 R R x Stenoplax purpurascens (C. B. Adams, 1845) C C C C C C C C x Stenoplax boogii (Haddon, 1886) R R R R x Subfamilia CALLISTOPLACINAE Género Callistochiton Carpenter in Dall, 1879 x Callistochiton shuttleworthianus Pilsbry, 1893 C C C C C C C C x Género Ceratozona Dall, 1882 x Ceratozona squalida (C. -

Gastropoda: Truncatellidae) from Florida, USA
THE NAUTILUS 130(1):13–16, 2016 Page 13 Three new species of Neogene Truncatella Risso, 1840 (Gastropoda: Truncatellidae) from Florida, USA Gary W. Schmelz Roger W. Portell 5575 Dogwood Way Florida Museum of Natural History Naples, FL 34116 University of Florida [email protected] 1659 Museum Road Gainesville, FL 32611 [email protected] ABSTRACT were discovered indicate that they were shallow water, marine environments. Three new species of truncatellid gastropods are described and illustrated. Two species are from the lower Miocene Chipola Formation of Calhoun County, Florida, and one is from upper Pliocene to lower Pleistocene deposits of the upper? Pinecrest MATERIALS AND METHODS beds, Tamiami Formation of Sarasota County, Florida. All three new species are believed to have inhabited shallow water, Specimens of Truncatella species from the lower marine environments. Miocene Chipola Formation were collected from two localities in the Florida Museum of Natural History, Invertebrate Paleontology Division (FLMNH IP) Direc- tory along the Chipola River in Calhoun County, Florida (Figure 1). One specimen was removed from matrix at FLMNH IP CA004, and two were collected from sub- INTRODUCTION merged deposits at FLMNH IP CA018. All sampled sediments were in situ. Furthermore, from the latter site, Gastropods of the family Truncatellidae display the ability pieces of loosely consolidated fossiliferous sandstone were to traverse vast distances of ocean to populate tropical and removed from underwater ledges with the aid of SCUBA. warm temperate environments. This ability to inhabit After removal, the material was brought to the FLMNH distant shorelines is accomplished by attaching them- IP laboratory for serial screen washing, air dried, and then selves or egg capsules to various pieces of flotsam and sediments were picked under stereomicroscope. -

Bulletin of the American Museum of Natural History Late Quaternary Fossil Mammals from the Cayman Islands, West Indies
LATE QUATERNARY FOSSIL MAMMALS FROM THE CAYMAN ISLANDS, WEST INDIES GARY S. MORGAN, ROSS D.E. MACPHEE, ROSEINA WOODS, AND SAMUEL T. TURVEY BULLETIN OF THE AMERICAN MUSEUM OF NATURAL HISTORY LATE QUATERNARY FOSSIL MAMMALS FROM THE CAYMAN ISLANDS, WEST INDIES GARY S. MORGAN New Mexico Museum of Natural History, Albuquerque, New Mexico ROSS D.E. MacPHEE Department of Mammalogy, American Museum of Natural History, New York ROSEINA WOODS School of Biological Sciences, Royal Holloway University of London Department of Earth Sciences, Natural History Museum, London SAMUEL T. TURVEY Institute of Zoology, Zoological Society of London, London BULLETIN OF THE AMERICAN MUSEUM OF NATURAL HISTORY Number 428, 79 pp., 24 figures, 8 tables Issued March 4, 2019 Copyright © American Museum of Natural History 2019 ISSN 0003-0090 CONTENTS Abstract.............................................................................3 Introduction.........................................................................3 Geographic and Geologic Setting.......................................................5 History of Paleontological Collecting in the Cayman Islands...............................7 Paleontological Materials and Methods ................................................10 Genomic Materials and Methods ......................................................10 Systematic Paleontology..............................................................12 Nesophontes Anthony, 1916 . 12 General Craniodental Features...................................................14