Microbial Carcinogenesis Lactic Acid Bacteria in Gastric Cancer
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Bacterial Dissimilation of Citric Acid Carl Robert Brewer Iowa State College
Iowa State University Capstones, Theses and Retrospective Theses and Dissertations Dissertations 1939 Bacterial dissimilation of citric acid Carl Robert Brewer Iowa State College Follow this and additional works at: https://lib.dr.iastate.edu/rtd Part of the Microbiology Commons Recommended Citation Brewer, Carl Robert, "Bacterial dissimilation of citric acid " (1939). Retrospective Theses and Dissertations. 13227. https://lib.dr.iastate.edu/rtd/13227 This Dissertation is brought to you for free and open access by the Iowa State University Capstones, Theses and Dissertations at Iowa State University Digital Repository. It has been accepted for inclusion in Retrospective Theses and Dissertations by an authorized administrator of Iowa State University Digital Repository. For more information, please contact [email protected]. INFORMATION TO USERS This manuscript has been reproduced from the microfilm master. UMI films the text directly from the original or copy submitted. Thus, some thesis and dissertation copies are in typewriter face, while others may be from any type of computer printer. The quality of this reproduction is dependent upon the quality of the copy submitted. Brol<en or indistinct print, colored or poor quality illustrations and photographs, print bleedthrough, substandard margins, and improper alignment can adversely affect reproduction. In the unlikely event that the author did not send UMI a complete manuscript and there are missing pages, these will be noted. Also, if unauthorized copyright material had to be removed, a note will indicate the deletion. Oversize materials (e.g., maps, drawings, charts) are reproduced by sectioning the original, beginning at the upper left-hand comer and continuing from left to right in equal sections with small overlaps. -

Citrobacter Koseri, Levinea Malonatica
INTERNATIONALJOURNAL OF SYSTEMATICBACTERIOLOGY, Jan. 1990, p. 107-108 Vol. 40, No. 1 0020-7713/90/010107-02$02.oo/o Copyright 0 1990, International Union of Microbiological Societies Correct Names of the Species Citrobacter koseri, Levinea malonatica , and Citrobacter diversus Request for an Opinion WILHELM FREDERIKSEN Department of Diagnostic Bacteriology and Antibiotics, Statens Seruminstitut, DK-2300 Copenhagen S,Denmark The single species carrying the three names Citrobacter koseri, Levinea malonatica, and Citrobacter diversus differs by at least eight characteristics from Citrobacter diversum as described by Werkman and Gillen in 1932. It is obviously not the same organism. Accordingly, the species should not carry the name proposed by Werkman and Gillen. I request that the name Citrobacter diversus be placed on the list of nomina rejicienda. Citrobacter koseri is the correct name. Levinea koseri is a correct combination when the genus Levinea is accepted. The epithet maZoonatica is a later synonym of the epithet koseri. In 1932 Werkman and Gillen (7) proposed a new genus, taxon C. diversum described by Werkman and Gillen in the Citrobacter, containing seven species. The description of following characteristics: motility, production of H,S, and Citrobacter diversum (sic) of Werkman and Gillen was based production of acid from inositol and raffinose. To this can be on two strains. These strains have not been kept available in added production of acid from glycogen, melizitose, starch, collections, and the name C. diversum did not come into and galactose, as Table 1 shows. general use. Ewing and Davis obviously accepted four deviations to In 1970 Frederiksen (4) described a new species which he lead to the statement about reactions “similar to those named Citrobacter koseri. -

Biomedical Applications of Bacteria-Derived Polymers
polymers Review Biomedical Applications of Bacteria-Derived Polymers Jonathan David Hinchliffe, Alakananda Parassini Madappura, Syed Mohammad Daniel Syed Mohamed and Ipsita Roy * Department of Materials Science and Engineering, Faculty of Engineering, University of Sheffield, Sheffield S1 3JD, UK; jhinchliffe3@sheffield.ac.uk (J.D.H.); [email protected] (A.P.M.); smdsyedmohamed1@sheffield.ac.uk (S.M.D.S.M.) * Correspondence: I.Roy@sheffield.ac.uk; Tel.: +44-11-4222-5962 Abstract: Plastics have found widespread use in the fields of cosmetic, engineering, and medical sciences due to their wide-ranging mechanical and physical properties, as well as suitability in biomedical applications. However, in the light of the environmental cost of further upscaling current methods of synthesizing many plastics, work has recently focused on the manufacture of these polymers using biological methods (often bacterial fermentation), which brings with them the advantages of both low temperature synthesis and a reduced reliance on potentially toxic and non-eco-friendly compounds. This can be seen as a boon in the biomaterials industry, where there is a need for highly bespoke, biocompatible, processable polymers with unique biological properties, for the regeneration and replacement of a large number of tissue types, following disease. However, barriers still remain to the mass-production of some of these polymers, necessitating new research. This review attempts a critical analysis of the contemporary literature concerning the use of a number of bacteria-derived polymers in the context of biomedical applications, including the biosynthetic Citation: Hinchliffe, J.D.; Parassini pathways and organisms involved, as well as the challenges surrounding their mass production. -

Carbapenem-Resistant Enterobacteriaceae a Microbiological Overview of (CRE) Carbapenem-Resistant Enterobacteriaceae
PREVENTION IN ACTION MY bugaboo Carbapenem-resistant Enterobacteriaceae A microbiological overview of (CRE) carbapenem-resistant Enterobacteriaceae. by Irena KennelEy, PhD, aPRN-BC, CIC This agar culture plate grew colonies of Enterobacter cloacae that were both characteristically rough and smooth in appearance. PHOTO COURTESY of CDC. GREETINGS, FELLOW INFECTION PREVENTIONISTS! THE SCIENCE OF infectious diseases involves hundreds of bac- (the “bug parade”). Too much information makes it difficult to teria, viruses, fungi, and protozoa. The amount of information tease out what is important and directly applicable to practice. available about microbial organisms poses a special problem This quarter’s My Bugaboo column will feature details on the CRE to infection preventionists. Obviously, the impact of microbial family of bacteria. The intention is to convey succinct information disease cannot be overstated. Traditionally the teaching of to busy infection preventionists for common etiologic agents of microbiology has been based mostly on memorization of facts healthcare-associated infections. 30 | SUMMER 2013 | Prevention MULTIDRUG-resistant GRAM-NEGative ROD ALert: After initial outbreaks in the northeastern U.S., CRE bacteria have THE CDC SAYS WE MUST ACT NOW! emerged in multiple species of Gram-negative rods worldwide. They Carbapenem-resistant Enterobacteriaceae (CRE) infections come have created significant clinical challenges for clinicians because they from bacteria normally found in a healthy person’s digestive tract. are not consistently identified by routine screening methods and are CRE bacteria have been associated with the use of medical devices highly drug-resistant, resulting in delays in effective treatment and a such as: intravenous catheters, ventilators, urinary catheters, and high rate of clinical failures. -

Citrobacter Koseri. II. Serological and Biochemical Examina- Tion of Citrobacter Koseri Strains from Clinical Specimens by B
J. Hyg., Camb. (1975), 75, 129 129 Printed in Great Britain Citrobacter koseri. II. Serological and biochemical examina- tion of Citrobacter koseri strains from clinical specimens BY B. ROWE, R. J. GROSS AND H. A. ALLEN Salmonella and Shigella Reference Laboratory, Central Public Health Laboratory, Colindale Avenue, London NW9 5HT (Received 30 January 1975) SUMMLARY 165 strains of Citrobacter koseri isolated from clinical specimens were studied and their biochemical reactions determined. They were examined serologically by means of a scheme consisting of 14 0 antigens. The sources of the clinical specimens were tabulated and the epidemiological information was summarized. The clinical significance of these findings is discussed. INTRODUCTION Bacterial strains variously described as Citrobacter koseri (Frederiksen, 1970), Citrobacter diversus (Ewing & Davis, 1972) and Levinea malonatica (Young, Kenton, Hobbs & Moody, 1971) should be regarded as members of a single species (Gross & Rowe, 1974). In this publication the name C. koseri has been used for convenience only, pending an agreement on the nomenclature of these organisms. In most genera within the family Enterobacteriaceae, serotyping has been invaluable for the precise identification of strains required in epidemiological investigations. An antigenic scheme for C. koseri has been described (Gross & Rowe, 1974) which initially consisted of seven 0 antigens and was subsequently expanded to 14 0 antigens (Gross & Rowe, 1975). The present study involves the examination of 165 strains from clinical sources. Their biochemical reactions and somatic 0 antigens have been determined and the clinical and epidemiological significance of these findings is discussed. MATERIALS AND METHODS Strains The sources of 165 strains isolated from clinical specimens are shown in Table 1. -

Antibiotic-Resistant Bacteria and Gut Microbiome Communities Associated with Wild-Caught Shrimp from the United States Versus Im
www.nature.com/scientificreports OPEN Antibiotic‑resistant bacteria and gut microbiome communities associated with wild‑caught shrimp from the United States versus imported farm‑raised retail shrimp Laxmi Sharma1, Ravinder Nagpal1, Charlene R. Jackson2, Dhruv Patel3 & Prashant Singh1* In the United States, farm‑raised shrimp accounts for ~ 80% of the market share. Farmed shrimp are cultivated as monoculture and are susceptible to infections. The aquaculture industry is dependent on the application of antibiotics for disease prevention, resulting in the selection of antibiotic‑ resistant bacteria. We aimed to characterize the prevalence of antibiotic‑resistant bacteria and gut microbiome communities in commercially available shrimp. Thirty‑one raw and cooked shrimp samples were purchased from supermarkets in Florida and Georgia (U.S.) between March–September 2019. The samples were processed for the isolation of antibiotic‑resistant bacteria, and isolates were characterized using an array of molecular and antibiotic susceptibility tests. Aerobic plate counts of the cooked samples (n = 13) varied from < 25 to 6.2 log CFU/g. Isolates obtained (n = 110) were spread across 18 genera, comprised of coliforms and opportunistic pathogens. Interestingly, isolates from cooked shrimp showed higher resistance towards chloramphenicol (18.6%) and tetracycline (20%), while those from raw shrimp exhibited low levels of resistance towards nalidixic acid (10%) and tetracycline (8.2%). Compared to wild‑caught shrimp, the imported farm‑raised shrimp harbored -

A Review of the Opportunistic Pathogen Citrobacter Freundii in Piglets Post Weaning : Public Health Importance
Sys Rev Pharm 2020;11(9):767-773 A multifaceted review journal in the field of pharmacy A Review of the Opportunistic Pathogen Citrobacter Freundii in Piglets Post Weaning : Public Health Importance Akvyan Rafi Hidayatullah1, Mustofa Helmi Effendi2*, Hani Plumeriastuti3, Freshindy Marissa Wibisono1, Erwan Budi Hartadi1, Eka Dian Sofiana1 1Master Program in Animal Diseases and Veterinary Public Health, Faculty of Veterinary Medicine, Universitas Airlangga. 2Departement of Veterinary Public Health, Faculty of Veterinary Medicine, Universitas Airlangga 3Departement of Veterinary Pathology, Faculty of Veterinary Medicine, Universitas Airlangga UNAIR C-Campus Mulyorejo, Surabaya, Indonesia, 60115. *Corresponding author: Mustofa Helmi Effendi. Department of Veterinary Public Health, Faculty of Veterinary Medicine, Universitas Airlangga, Surabaya, Indonesia, Postal code: 60115. Email: [email protected] ABSTRACT The review study will describe clearly related research about Citrobacter Keywords: Citrobacter freundii, microbiota, piglets, antimicrobial freundii in piglet intestines which has been known to play a very important resistance, public health role in animal health and its effects on aspects of public health. Thus, the existence of Citrobacter freundii as a pathogenic opportunistic bacterium as a Correspondence: result of weaning plays an important role in the formation and maintenance Mustofa Helmi Effendi of the intestinal microbiota which affects the health and growth performance Department of Veterinary Public Health, Faculty of Veterinary -

Ecology of Pathogenic E.Coli - Stefano Morabito
MEDICAL SCIENCES - Ecology Of Pathogenic E.Coli - Stefano Morabito ECOLOGY OF PATHOGENIC E. coli Stefano Morabito European Reference Laboratory for Escherichia coli including VTEC. Foodborne zoonoses Unit; Department of Veterinary Public Health and Food Safety Istituto Superiore di Sanità, Viale Regina Elena 299, 00161, Rome. Italy. Keywords: Escherichia coli, pathogenicity, genomic plasticity, survival, environment, colonization, animal reservoirs, zoonosis. Contents 1. Introduction 2. Pathogenic E. coli types 3. Ecology of Pathogenic E. coli Circulating in Humans 4. The Zoonotic Connection 5. Ecology of Zoonotic E. coli 6. Pathogenic Escherichia coli Producing Biofilm 7. Signaling In the Gastro-Intestinal Tract and Pathogenesis 8. E. coli Pathotypes with Atypical Combination of Virulence Factors 9. Acknowledgments Glossary Bibliography Biographical Sketch Summary Escherichia coli is a ubiquitous bacterial species. It is part of the intestinal microflora of humans and warm-blooded animals and, at the same time, is among the most diffuse bacterial species in the environment. The pivotal aspect of this success is represented by an exceptional genomic plasticity. As a matter of fact, E. coli is capable to exchange genetic material with other bacteria, often belonging to different species, through horizontal gene transfer, a process involving the action of mobile genetic elements, such as bacteriophages, transposons, pathogenicity islands and plasmids. The relationships between E. coli and the human host are generally based on commensalism and yet this species exerts a beneficial effect in the gut, however, some strains evolved the capability to harm and cause disease either involving the gastro- intestinal tract or in other organism‘s districts. Most of the pathogenic E. -

Bacterial Biosurfactant from Citrobacter Freundii MG812314.1 As a Bioremoval Tool of Heavy Metals from Wastewater Eman Z
Gomaa and El-Meihy Bulletin of the National Research Centre (2019) 43:69 Bulletin of the National https://doi.org/10.1186/s42269-019-0088-8 Research Centre RESEARCH Open Access Bacterial biosurfactant from Citrobacter freundii MG812314.1 as a bioremoval tool of heavy metals from wastewater Eman Z. Gomaa1* and Rasha M. El-Meihy2,3 Abstract Background: The problem of heavy metal contamination is a global issue, and the challenge is to develop methods to remove heavy metals from soil and water. Recently, biosurfactants are one of the compounds that provide an attractive eco-friendly alternative to the physicochemical process in alleviating the heavy metals. Results: Sixty bacterial isolates were isolated from Al-Rahawy drain sediments and screened for biosurfactant production. Only 10 isolates were recoded as biosurfactant producers by the oil spreading and emulsifying assays in addition to the ability of biosurfactant on heavy metal removal in wastewater. The most potent isolate was identified using morphological, cultural, biochemical characteristics, antibiotics susceptibility, and 16 s rRNA technique as Citrobacter freundii MG812314.1. Conclusions: The produced biosurfactant was found to be more effective in removing heavy metals from wastewater, viz 80, 67, 66, 55, 45, 44, and 41% of aluminum, lead, zinc, cadmium, iron, copper, and manganese, respectively, under two inoculum potentials and two contact time. The interaction of heavy metals with biosurfactant was monitored using scanning electron microscope (SEM), energy dispersive X-ray spectra (EDX), and Fourier-transform infrared spectroscopy (FTIR) analyses. Keywords: Heavy metals, Biosurfactant, Wastewater, Bioremediation, Citrobacter freundii Introduction (Ezzat et al. 2012). These metals cause serious ecological Biosphere pollution due to heavy metals is a confused problems because they are non-degradable and can bio- problem that causes negative effects on various environ- accumulate through the food chain (Goher et al. -

Activities and Prevalence of Proteobacteria Members Colonizing Echinacea Purpurea Fully Account for Macro- Phage Activation Exhibited by Extracts of This Botanical
1258 Original Papers Activities and Prevalence of Proteobacteria Members Colonizing Echinacea purpurea Fully Account for Macro- phage Activation Exhibited by Extracts of This Botanical Authors Mona H. Haron 1*, Heather L. Tyler 2, 3*, Nirmal D. Pugh 1, Rita M. Moraes1, Victor L. Maddox4, Colin R. Jackson 2, David S. Pasco1,5 Affiliations The affiliations are listed at the end of the article Key words Abstract highest potency for in vitro macrophage activa- l" Echinacea purpurea ! tion and were the most predominant taxa. Fur- l" Asteraceae Evidence supports the theory that bacterial com- thermore, the mean activity exhibited by the l" immunostimulatory munities colonizing Echinacea purpurea contrib- Echinacea extracts could be solely accounted for l" macrophage activation ute to the innate immune enhancing activity of by the activities and prevalence of Proteobacteria l" bacteria l" proteobacteria this botanical. Previously, we reported that only members comprising the plant-associated bacte- l" endophytes about half of the variation in in vitro monocyte rial community. The efficacy of E. purpurea mate- stimulating activity exhibited by E. purpurea ex- rial for use against respiratory infections may be tracts could be accounted for by total bacterial determined by the Proteobacterial community load within the plant material. In the current composition of this plant, since ingestion of bac- study, we test the hypothesis that the type of bac- teria (probiotics) is reported to have a protective teria, in addition to bacterial load, is necessary to effect against this health condition. fully account for extract activity. Bacterial com- munity composition within commercial and freshly harvested (wild and cultivated) E. -
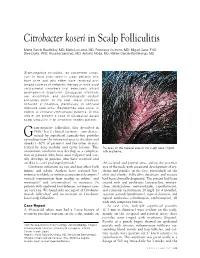
Citrobacter Koseri in Scalp Folliculitis
Citrobacter koseri in Scalp Folliculitis Marta Garcia-Bustinduy, MD; Maria Lecuona, MD; Francisco Guimera, MD; Miguel Saez, PhD; Sara Dorta, PhD; Rosalba Sanchez, MD; Antonio Noda, MD; Rafael Garcia-Montelongo, MD Gram-negative folliculitis, an uncommon condi- tion, is most often seen in older patients who have acne and who either have received pro- longed courses of antibiotic therapy or have used antibacterial cleansers that selectively inhibit gram-positive organisms. Citrobacter infections are uncommon, and dermatologists seldom encounter them. In the past, these infections occurred in hospitals, particularly in neonatal intensive care units. Bacteremias also occur in elderly or immunocompromised patients. In this article, we present a case of Citrobacter koseri scalp folliculitis in an otherwise healthy patient. ram-negative folliculitis, first described in 1968,1 has 2 clinical varieties—one charac- G terized by superficial comedo-free pustules extending from the infranasal area to the chin and cheeks (~80% of patients) and the other charac- terized by deep nodular and cystic lesions. This Pustules on the parietal area of the scalp were ringed uncommon condition may develop as a complica- with erythema. tion in patients who have acne vulgaris2 and usu- ally develops in patients who have received oral antibiotics over prolonged periods.3 the occipital and parietal areas, and on the posterior Citrobacter infections are rare and may affect both area of the neck, with occasional development of ery- infants and adults. Authors have reported bac- thema and papules on the face, particularly on the teremias in elderly or immunocompromised patients,4 chin and cheeks. Folliculitis decalvans and rosacea vertical transmission from mother to infant,5 and had been clinically diagnosed. -

Citrobacter Freundii
P.O. Box 131375, Bryanston, 2074 Ground Floor, Block 5 Bryanston Gate, 170 Curzon Road Bryanston, Johannesburg, South Africa 804 Flatrock, Buiten Street, Cape Town, 8001 www.thistle.co.za Tel: +27 (011) 463 3260 Fax: +27 (011) 463 3036 Fax to Email: + 27 (0) 86‐557‐2232 e‐mail : [email protected] Please read this section first The HPCSA and the Med Tech Society have confirmed that this clinical case study, plus your routine review of your EQA reports from Thistle QA, should be documented as a “Journal Club” activity. This means that you must record those attending for CEU purposes. Thistle will not issue a certificate to cover these activities, nor send out “correct” answers to the CEU questions at the end of this case study. The Thistle QA CEU No is: MT-13/00142. Each attendee should claim THREE CEU points for completing this Quality Control Journal Club exercise, and retain a copy of the relevant Thistle QA Participation Certificate as proof of registration on a Thistle QA EQA. MICROBIOLOGY LEGEND CYCLE 33 ORGANISM 5 Citrobacter freundii Citrobacter freundii are facultative anaerobic Gram-negative coliform bacteria in the Enterobacteriaceae family. The bacteria are long rod-shaped with a typical length of 1-5 μm. Most C. freundii cells are surrounded by several flagella used for locomotion, but a few are non-motile. Citrobacter species are differentiated by their ability to convert tryptophan to indole, ferment lactose, and use malonate. C. freundii can use citrate as a sole carbon source. The Citrobacter genus was discovered in 1932 by Werkman and Gillen.