Genetic Evaluations of the German Sport Horse: Population Structure and Use of Data from Foal and Mare Inspections and Performance Tests of Mares
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
August 13-16 Purchase Tickets at Hayesco.Ca - Use Code Ctjump for 10% Off 4 Days • 40 Vendors • Live Music Kidzone • Top Show Jumping
AUGUST 13-16 PURCHASE TICKETS AT HAYESCO.CA - USE CODE CTJUMP FOR 10% OFF 4 DAYS • 40 VENDORS • LIVE MUSIC KIDZONE • TOP SHOW JUMPING Mailing Registration No. 40032702 CaledonCitizenCombined Thursday Circulation: 36,816 │ 905-857-6626 │ 1-888-557-6626 │ www.caledoncitizen.com $1.00 per copy (95¢ + 5¢ G.S.T.) Thursday, July 30, 2015 Volume 34, Issue 30 THE Kostynyk Denture Centre DENTURIST | DENTURE CENTRE | SINCE 1995 Loose Dentures? We can help Emergency Repairs while you wait. To Schedule Your Free Consultation Please Call David Kostynyk, DD, BSc. 905-857-4464 30 Martha St., Suite 105 Signs like this one will soon be marking Emil Kolb Parkway. Joining Kolb for Tuesday;s celebration were Brampton Councillor John Sprovieri, Caledon Councillor Rob Mezzapelli, Peel Regional Chair Frank Dale, Mayor Allan Thompson and Caledon Councillor Nick deBoer. Photo by Bill Rea Emil Kolb Parkway to open soon Sales • Installation By Bill Rea “The project is well overdue, sure off the valley. returned, several councillors Service • Rentals It’s been talked about for de- and will be a tremendous ben- Kolb said he convinced were laughing. Delivering Superior Service Since 1990 cades, but the fi nal link in the efi t for the community,” Dale Town council in 1983 to start “I guess we should tell Emil,” Bolton bypass is about to open. commented. a full environmental assess- he remembered Mississauga 905-586-COOL (2665) The entire road itself is not “We have certainly named ment on a possible bypass, Councillor Pat Saito saying. slated to open until Aug. 10, the road appropriately,” he with the study area stretch- “It’s truly an honour to have donsheatingandcooling.com but there was a celebration of added. -

List of Horse Breeds 1 List of Horse Breeds
List of horse breeds 1 List of horse breeds This page is a list of horse and pony breeds, and also includes terms used to describe types of horse that are not breeds but are commonly mistaken for breeds. While there is no scientifically accepted definition of the term "breed,"[1] a breed is defined generally as having distinct true-breeding characteristics over a number of generations; its members may be called "purebred". In most cases, bloodlines of horse breeds are recorded with a breed registry. However, in horses, the concept is somewhat flexible, as open stud books are created for developing horse breeds that are not yet fully true-breeding. Registries also are considered the authority as to whether a given breed is listed as Light or saddle horse breeds a "horse" or a "pony". There are also a number of "color breed", sport horse, and gaited horse registries for horses with various phenotypes or other traits, which admit any animal fitting a given set of physical characteristics, even if there is little or no evidence of the trait being a true-breeding characteristic. Other recording entities or specialty organizations may recognize horses from multiple breeds, thus, for the purposes of this article, such animals are classified as a "type" rather than a "breed". The breeds and types listed here are those that already have a Wikipedia article. For a more extensive list, see the List of all horse breeds in DAD-IS. Heavy or draft horse breeds For additional information, see horse breed, horse breeding and the individual articles listed below. -

Ocala Jockey Club Breeding Info
Ocala Jockey Club HT Breeding Info Rider Last Name Rider First name Horse Sire Dam Dam Sire Breed Owner Breeder Division Sex Aden Nicole Truckee Bash Truckee xx Bashful Belle xx Stately Cielo xx Thoroughbred Woods Baughman Four Quarters Corp CCI4-S Gelding (CA) Aharoni Arielle Dutch Times Good Times Alino Queen Michelino Dutch Warmblood Christina Aharoni Lauren Efford CCI4-L Gelding Atkinson James Fleur de Lis Heartbreaker Matana U Hattrick Dutch Warmblood James Atkinson. Griendstveen HJJJ CCII3-L Gelding Jim Seilsopour Bouwmans Babbitt Charlotte 2 A.M. Sheraton Regina-K Ahorn Dutch Warmblood Charlotte Babbitt A. Lusseveld CCI3-L Gelding Baker Shanon Ballingowan Zeal Seabrook Shara Bride Clover Hill Irish Sport Horse Shannon Baker Tom Reilly CCI3-L Gelding Baugh Alexandera I Spye Harlequin Du Carel Lombardos Corner Lombardo Irish Sport Horse. Jesse Campbell Thomas Ryan CCI2-L Gelding Baugh Alexandra Mr. Candyman Canto 16 Montara Corofino I Holsteiner Altorac Farm Meerheimb H.W. CCI4-L Gelding Freiherr Von Beshear Emily El Mesano Mizzen Mast La Laja El Prado Thoroughbred Emily Beshear Helen K. Groves CCI2-L Gelding Revokable Trust Beshear Emily Deal With It Medaglia d’Oro Amada Unbridled Thoroughbred Emily Beshear CCI3-L Gelding Beshear Emily Templewood Horse Chestnut Missy Dear Deerhound Thoroughbred Catherine Birley Kenneth Tomlinson CCI2-L Gelding Black Maya Fe Chardonnay Clinton I Callina Compliment Hanoverian Light Speed CCI2-L Gelding Equestrian, LLC Black Maya Maks Mojo C Mighty Magic Winter Morning Ramiro’s Bube Hanoverian Laurie Cameron Laurie Cameron CCI2-L Gelding Bowman Sarah Altus Louvo Quitus Louvo Vanille du Tertre J’T’Adore Selle Francais Sarah Bowman M. -

Electronic Supplementary Material - Appendices
1 Electronic Supplementary Material - Appendices 2 Appendix 1. Full breed list, listed alphabetically. Breeds searched (* denotes those identified with inherited disorders) # Breed # Breed # Breed # Breed 1 Ab Abyssinian 31 BF Black Forest 61 Dul Dülmen Pony 91 HP Highland Pony* 2 Ak Akhal Teke 32 Boe Boer 62 DD Dutch Draft 92 Hok Hokkaido 3 Al Albanian 33 Bre Breton* 63 DW Dutch Warmblood 93 Hol Holsteiner* 4 Alt Altai 34 Buc Buckskin 64 EB East Bulgarian 94 Huc Hucul 5 ACD American Cream Draft 35 Bud Budyonny 65 Egy Egyptian 95 HW Hungarian Warmblood 6 ACW American Creme and White 36 By Byelorussian Harness 66 EP Eriskay Pony 96 Ice Icelandic* 7 AWP American Walking Pony 37 Cam Camargue* 67 EN Estonian Native 97 Io Iomud 8 And Andalusian* 38 Camp Campolina 68 ExP Exmoor Pony 98 ID Irish Draught 9 Anv Andravida 39 Can Canadian 69 Fae Faeroes Pony 99 Jin Jinzhou 10 A-K Anglo-Kabarda 40 Car Carthusian 70 Fa Falabella* 100 Jut Jutland 11 Ap Appaloosa* 41 Cas Caspian 71 FP Fell Pony* 101 Kab Kabarda 12 Arp Araappaloosa 42 Cay Cayuse 72 Fin Finnhorse* 102 Kar Karabair 13 A Arabian / Arab* 43 Ch Cheju 73 Fl Fleuve 103 Kara Karabakh 14 Ard Ardennes 44 CC Chilean Corralero 74 Fo Fouta 104 Kaz Kazakh 15 AC Argentine Criollo 45 CP Chincoteague Pony 75 Fr Frederiksborg 105 KPB Kerry Bog Pony 16 Ast Asturian 46 CB Cleveland Bay 76 Fb Freiberger* 106 KM Kiger Mustang 17 AB Australian Brumby 47 Cly Clydesdale* 77 FS French Saddlebred 107 KP Kirdi Pony 18 ASH Australian Stock Horse 48 CN Cob Normand* 78 FT French Trotter 108 KF Kisber Felver 19 Az Azteca -

BWP/NAD FOAL REGISTRATION PACKET for Foals Not Attending a Keuring
BWP/NAD FOAL REGISTRATION PACKET For Foals Not Attending a Keuring ! Dear Mare Owner; Congratulations on the birth of your foal! We know this is an exciting time for you and we want to make the registration process as seamless as possible. To qualify for registration with out inspection, both sire and dam of the foal must be approved with the BWP or BWP/NAD. You may attend a future keuring for branding up to the age of two and also enter yearling & 2YO futurities for inspection. Contact [email protected] if you have any questions about your registration options. Please read the instructions carefully and be sure to provide all the required documents by November 15th of the foaling year to the following address: BWP/NAD, Attn: Chris Sallee, 41 York Drive, St. Louis, MO 63144 or email to [email protected] You will need to provide the following documents: 1. DECLARATION OF BIRTH Be sure to sign in the spaces provided at the bottom of the form. The Breeder is the owner or lessee of the mare at the time of foaling. 2. MARKINGS DIAGRAM Please have a veterinarian complete the markings diagram. White markings are to be outlined in RED ink. Whorls, scars, brands and other markings should be noted in BLACK. Please note the whorls on the foals face, poll region, along the under side of the neck or throat and each side of the crest with an ‘X’. 3. BREEDING CERTIFICATE You must provide a breeding certificate from the stallion owner or contract for the purchase of semen and/or semen the bill of sale. -

An Analysis of the Inbreeding Level of Holstein Stallions Presented for Classification in 2003–2012
Acta Sci. Pol. Zootechnica 16(2) 2017, 35–38 www.asp.zut.edu.pl pISSN 1644-0714 eISSN 2300-6145 DOI:10.21005/asp.2017.16.2.05 Received: 24.05.2017 ORIGINAL PAPER Accepted: 17.08.2017 AN ANALYSIS OF THE INBREEDING LEVEL OF HOLSTEIN STALLIONS PRESENTED FOR CLASSIFICATION IN 2003–2012 Ryszard Pikuła1B, Wilhelm Grzesiak2, Daniel Zaborski2, Wioletta Werkowska1 1Laboratory of Horse Breeding and Animal Assisted Therapy, West Pomeranian University of Technology, Szczecin, Doktora Judyma 14, 71-466 Szczecin, Poland 2Department of Ruminants Science, West Pomeranian University of Technology, Szczecin, Klemensa Janickiego 29, 71-466 Szczecin, Poland ABSTRACT The aim of the present study was to analyze and compare the inbreeding level of the Holstein stallions presented for classification for breeding in 2003–2012 in Germany. The data were collected from the Holsteiner Körung und Reitepferde – Auktion catalogues. Based on the analysis of 869 pedigrees (up to the fifth generation inclusive), inbreeding coefficients (F), expressed as a percentage for each stallion, were calculated. The highest F values were recorded in 2003 (2.05%) and 2004 (1.74%). Statistically significant differences in the F values of the stallions from individual years were found. The outcomes of the present study show that the breeding work based on conscious assumptions is carried out in order to increase an inbreeding level in the sport horses’ production, regarding at the same time that the value of the inbreeding coefficient cannot be too high. Key words: inbreeding, Holstein, stallions, classification, breeding INTRODUCTION standing ancestors and subsequently consolidating desi- red traits [Pikuła and Werkowska 2009, 2011a, 2011b]. -

$25,000 American Standard Markel Insurance Grand Prix Saturday August 1, 2020 # HORSE BREED RIDER OWNER SCORE
$25,000 American Standard Markel Insurance Grand Prix Saturday August 1, 2020 # HORSE BREED RIDER OWNER SCORE 1 1541 SCARFACE CZECH, 2009, BAY, Mare, ARISTO Z x SANTA ALEXIS SOKOLOV ALEXIS SOKOLOV 2 1439 FOOTLOOSE KWPN, 2010, BAY, Mare, BERLIN x LEGATA MARI GROMKOWSKI CALIBER SHOW JUMPERS LLC WESTPHALIAN, 2009, BROWN, Mare, ARIOSO DU THEILLET x 3 1131 ARIELL LA SIRENE STELLA KERI POTTER HANNAH LOLY UNKNOWN, 2006, GREY, Mare, CARDENTO x NARCOTIQUE DU 4 1133 AYMA DE LA DEMI LUNE HOUSSOIT HANNAH LOLY HANNAH LOLY SELLE FRANCAIS, 2013, BAY, Mare, EL DORADO VAN DE 5 1662 GOOD LUCK ZESHOEK x UFENIA SIMON SCHROEDER MORAD ALMASRI SELLE FRANCAIS, 2008, BROWN, Stallion, NIAGARA D'ELLE x 6 1596 UNIQUE STAR CASSANDRA D'ICK HANNA MAURITZSON RITZ FUENTE LLC 7 946 CHARDONNAY RPSI, 2004, BAY, Mare, CON NIOR x LILLY ROBERT BLANCHETTE ROBERT BLANCHETTE ZANGERSHEIDE, 2010, BAY, Gelding, CHELLANO Z x CADENCE 8 746 CIRUS DU RUISSEAU Z DU RUISSEAU Z ZUME GALLAHER BLUE GATE STABLES, LLC 9 747 CONCOLUE WARMBLOOD, 2010, BAY, Gelding, CONTHARGOS x DUGATRIA NICOLE HAUNERT CHEROKEE SHOW HORSES INC 10 360 CORNET WURTTEMBERGER, 2010, BAY, Gelding, COLESTUS x MARAICA CASSIO RIVETTI NEIL JONES EQUESTRIAN, INC BELGIAN WARMBLOOD, 2009, BAY, Gelding, OGANO SITTE x 11 1317 JARPUR FIEBE VAN'T WATERLATJE LINDSAY ARCHER RHYS FARMS, LLC BELGIAN WARMBLOOD, 2011, BAY, Gelding, BAMAKO DE MUZE 12 1318 LUIGI VD BISSCHOP x EVITA VAN'T ROOSAKKER MATT ARCHER RHYS FARMS, LLC HOLSTEINER WARMBLOOD, 2005, DARK BROWN, Stallion, 13 1466 CHANTICO CONTENDER x WIENA JOHN PEARCE KAREN BALL 14 -

Der Hannoveraner Eng
10|2020 DER HANNOVERANER No. 10 | October 2020 Sport Hannover‘s trump cards Hannoveraner Verband The course is newly set Bundeschampion of three-year old mares and geldings: Rock Festival by Rock Fore- ver/Fürstenball and rider Joline Durand. Sport Bundeschampion of three-year old stallions: Va’Pensiero by Vitalis/Fürstenball and Hannah Laser. Photo: Equitaris Hannover‘s trump cards Hannoveraner horses won four medals – two times gold and two times silver – in four riding horse classes at the Bundeschampionate in September. Hannover’s trump cards stood out especially in both classes for three-year olds. By Dr. Ludwig Christmann 2 Der Hannoveraner 10|2020 Sport he final competition at the Bundeschampion- was bred in Denmark by Sally Hjort Schultz. He too Tate was the class for three-year old stallions. impressed with his rideability, his character and his From a Hannoveraner perspective, this was the ab- performance willingness. With respect to his type, solute highlight. For the first time, the final for his large frame immediately catches one’s eye, three-year olds was scheduled without a guestrid- which is paired with his strong type and his long- er test. Va’Pensiero by Vitalis/Fürstenball out of the legs. His rider Sina Aringer brilliantly developed his breeding program of Breeding Farm Düvel in uphill movement, which reflected high elasticity. Katlenburg became the winner. Va’Pensiero is a The Westphalian stallion Macchiato by Morricone/ very special horse. In addition to his qualities as a Just Perfect (owner: Stallion Station Pape) earned rising dressage horse, he impresses with his dispo- bronze (final score 9.0) with rider Greta Heemsoth, sition and his superior demeanor. -

Breed Societies and Breeding Operations Based in Other
Date of last update: 23/08/2021 BREED SOCIETIES AND BREEDING OPERATIONS BASED IN OTHER EU MEMBER STATES WHICH HAVE OFFICIALLY NOTIFIED THEIR ACTIVITIES IN THE REPUBLIC OF IRELAND IN LINE WITH ARTICLE 12 AND ARTICLE 64(6) OF REGULATION (EU) 2016/1012 Name of Member State Species Name and Contact Name of the where the breeding Detail Breed organization is recognised Denmark Porcine Landbrug & Fødevarer Dansk Landrace (DanAvl) Danish Agriculture & Food Dansk Yorkshore Council (DanAvl) Dansk Duroc Axeltorv 3 DK 1609 København V +45 3339 4000 [email protected] https://www.lf.dk/ Denmark Hybrid Breeding Pigs Landbrug & Fødevarer DanBred Hybrid (DanAvl) Danish Agriculture (based on Dansk & Food Landrace, Dansk Council (DanAvl) Yorkshire, and Dansk Axeltorv 3 Duroc) DK 1609 København V +45 3339 4000 [email protected] https://www.lf.dk/ Germany Equine Deutsche Quarter Horse Association e.V. American Quarter https://tgrdeu.genres.de/veroeffentlichungvoninformationengema Horse esstierzuchtrecht/zuechtervereinigungen Deutsche Quarter Horse Association e.V. Daimlerstrasse 22 63741 Aschaffenburg phone:+49 (0) 6021 58459 0 Fax:+49 (0) 6021 58459 79 E-Mail: info@)dqha.de Internet: http://www.dqha.de Germany Equine Westfälisches Pferdestammbuch e.V. Westfälisches Reitpferd https://tgrdeu.genres.de/veroeffentlichungvoninformationengema esstierzuchtrecht/zuechtervereinigungen Westfälisches Pferdestammbuch e.V. Sudmühlenstraße 33 48157 Münster-Handorf phone: +49 (0) 251 32809 0 Fax: +49 (0) 251 32809 24 E-Mail: [email protected] Internet: http://www.westfalenpferde.de Germany Equine Verband der Züchter des Holsteiner Pferdes e.V. Holsteiner Warmblut https://tgrdeu.genres.de/veroeffentlichungvoninformationenge maesstierzuchtrecht/zuechtervereinigungen Verband der Züchter des Holsteiner Pferdes e.V. Steenbeker Weg 151 24106 Kiel phone: +49 (0) 431 3059960 Fax:+49 (0) 431 336142 E-Mail: [email protected] Internet: http://www.holsteiner-verband.de Germany Equine Springpferdezuchtverband Oldenburg International e.V. -
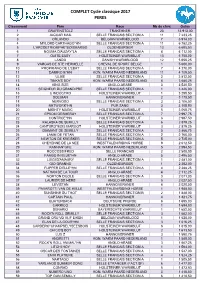
2017 Complet Cycle Classique
COMPLET Cycle classique 2017 PERES Classement Père Race Nb de chvx Gains 1 GRAFENSTOLZ TRAKEHNER 23 13 913,00 2 JAGUAR MAIL SELLE FRANCAIS SECTION A 11 7 233,25 3 ORLANDO BELGIAN WARMBLOOD 7 6 914,00 4 MYLORD CARTHAGO*HN SELLE FRANCAIS SECTION A 11 6 782,25 5 L'ARCDETRIOMPHE*BOISMARGO OLDENBURGER 13 6 685,50 6 NOUMA D'AUZAY*LA SELLE FRANCAIS SECTION A 8 6 112,00 7 QUITE EASY HOLSTEINER WARMBLUT 13 5 758,00 8 LANDO DANISH WARMBLOOD 12 5 656,25 9 VARGAS DE STE HERMELLE CHEVAL DE SPORT BELGE 1 5 600,00 10 PROPRIANO DE L'EBAT SELLE FRANCAIS SECTION A 7 4 270,00 11 DAMIRO B*HN KON. WARM PAARD NEDERLAND 11 4 109,50 12 ULIXE SELLE FRANCAIS SECTION A 3 3 812,50 13 TINKA'S BOY KON. WARM PAARD NEDERLAND 8 3 680,25 14 KING SIZE ANGLO-ARABE 3 3 530,50 15 SEIGNEUR DU GRAND PRE SELLE FRANCAIS SECTION A 1 3 426,00 16 C INDOCTRO HOLSTEINER WARMBLUT 1 3 299,50 17 SOLIMAN HANNOVERANER 2 3 179,50 18 NERVOSO SELLE FRANCAIS SECTION A 3 3 106,50 19 NETWORK*HN PUR SANG 3 3 105,50 20 MIGHTY MAGIC HOLSTEINER WARMBLUT 5 3 068,75 21 EPSOM GESMERAY SELLE FRANCAIS SECTION A 7 2 997,75 22 CONTROE*HN HOLSTEINER WARMBLUT 6 2 987,50 23 KALASKA DE SEMILLY SELLE FRANCAIS SECTION A 6 2 979,25 24 CANTURO*BOIS MARGOT HOLSTEINER WARMBLUT 3 2 879,25 25 DIAMANT DE SEMILLY SELLE FRANCAIS SECTION A 5 2 866,75 26 LAMM DE FETAN SELLE FRANCAIS SECTION A 2 2 760,00 27 NEWTON DE KREISKER SELLE FRANCAIS SECTION A 5 2 725,00 28 CHEYENNE DE LA NEE WESTFALEN RIDING HORSE 9 2 612,50 29 KANNAN*GFE KON. -

International Jumping Festival Grand Prix Field Wednesday, September 18, 2019
INTERNATIONAL JUMPING FESTIVAL GRAND PRIX FIELD WEDNESDAY, SEPTEMBER 18, 2019 *Horses showing in the USEF Talent Search Finals may only show in class #229 1.15m Jumpers on Wednesday. 8:00 a.m. – Classes #221. 1.00m Jumpers Table II, Sec. 2(d) 11:00 a.m. – Classes #229. 1.15m Jumpers Table II, Sec. 2(d)* *Open water will be an option 1:00 p.m. – Class #250. 1.20m Jumpers Table II, Sec. 2(d) 3:15 p.m. – Class #305. 1.40m Jumpers Table II, Sec. 1 Cancelled – Class #309. 1.45m Jumpers Table II, Sec. 1 [Finish: 3:50 p.m.] Class #221 - 1.00m Jumpers # HORSE RIDER ELVIS 284 HOLSTEINER, 2010, G, 1 542 LARIMAR x WINTERA JAVIER ABAD 8:00:00 CANTINA TRAKEHNER, 2013, G, 2 262 UNKNOWN x UNKNOWN JOHN BRAGG 8:02:30 BOUJEE SELLE FRANCAIS, 2010, G, 3 464 UNKNOWN x UNKNOWN JOSH MADGWICK 8:05:00 CAMPITELLO 5 HANOVERIAN, 2007, G, 4 581 CATOKI x GINA EMMA CATHERINE REICHOW 8:07:30 NEWTON HOLSTEINER, 2013, G, 5 326 NARRADO x SEESTERMUEHE JENNY KARAZISSIS 8:10:00 KANNAN LOVER WARMBLOOD, 2002, G, 6 232 KANNAN x PENNY LOVER EMILY ESAU WILLIAMS 8:12:30 MLB CLAIR DE LUNE UNKNOWN, 2013, M, 7 379 INDOCTRO x ALLURE MICHELLE PARKER 8:15:00 CAZZ C DUTCH WARMBLOOD, 2007, 8 136 G, UNKNOWN x UNKNOWN JACKIE LEFAVE 8:17:30 DEBUT HANOVERIAN, 2011, G, 9 532 DAMSEY x GENOVEVA LEXI WEDEMEYER 8:20:00 ANAKIN DUTCH WARMBLOOD, 2012, G, VAN HELSING x DYTHERA 10 295 H BROOKE MOSTMAN 8:22:30 S & L TECHNICOLOR DUTCH WARMBLOOD, 2006, 11 393 G, CHIN CHIN x UNKNOWN ALYSIA LYNCH-SHERARD 8:25:00 HANDS UP DUTCH WARMBLOOD, 2012, 12 383 G, INDOCTRO x SUZANNA LAUREN KATZENELLENBOGEN 8:27:30 -

Breeding Programme for Oldenburg Horses
Breeding Programme for Oldenburg Horses Verband der Züchter des Oldenburger Pferdes e.V. Grafenhorststr. 5 · 49377 Vechta · Germany Telephone: 04441 -9355 -0 Fax: 04441 -9355 -99 [email protected] www.oldenburger-pferde.com Breeding Programme for Oldenburg Horses Breeding Programme for Oldenburg Horses 1. Information on the Studbook.....................................................................................................3 2. Geographical area ....................................................................................................................3 3. Size of the breeding population ................................................................................................3 4. Breeding goal ...........................................................................................................................3 5. Characteristic features and qualities .........................................................................................3 5.1. Exterior .....................................................................................................................................3 5.2. Movement incl. jumping ............................................................................................................4 5.3 Interior qualities .........................................................................................................................5 6. Selection characteristics ...........................................................................................................5