Endangered Wild Salmon Infected by Newly Discovered Viruses
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Southwest Guangdong, 28 April to 7 May 1998
Report of Rapid Biodiversity Assessments at Qixingkeng Nature Reserve, Southwest Guangdong, 29 April to 1 May and 24 November to 1 December, 1998 Kadoorie Farm and Botanic Garden in collaboration with Guangdong Provincial Forestry Department South China Institute of Botany South China Agricultural University South China Normal University Xinyang Teachers’ College January 2002 South China Biodiversity Survey Report Series: No. 4 (Online Simplified Version) Report of Rapid Biodiversity Assessments at Qixingkeng Nature Reserve, Southwest Guangdong, 29 April to 1 May and 24 November to 1 December, 1998 Editors John R. Fellowes, Michael W.N. Lau, Billy C.H. Hau, Ng Sai-Chit and Bosco P.L. Chan Contributors Kadoorie Farm and Botanic Garden: Bosco P.L. Chan (BC) Lawrence K.C. Chau (LC) John R. Fellowes (JRF) Billy C.H. Hau (BH) Michael W.N. Lau (ML) Lee Kwok Shing (LKS) Ng Sai-Chit (NSC) Graham T. Reels (GTR) Gloria L.P. Siu (GS) South China Institute of Botany: Chen Binghui (CBH) Deng Yunfei (DYF) Wang Ruijiang (WRJ) South China Agricultural University: Xiao Mianyuan (XMY) South China Normal University: Chen Xianglin (CXL) Li Zhenchang (LZC) Xinyang Teachers’ College: Li Hongjing (LHJ) Voluntary consultants: Guillaume de Rougemont (GDR) Keith Wilson (KW) Background The present report details the findings of two field trips in Southwest Guangdong by members of Kadoorie Farm & Botanic Garden (KFBG) in Hong Kong and their colleagues, as part of KFBG's South China Biodiversity Conservation Programme. The overall aim of the programme is to minimise the loss of forest biodiversity in the region, and the emphasis in the first three years is on gathering up-to-date information on the distribution and status of fauna and flora. -

2020 Taxonomic Update for Phylum Negarnaviricota (Riboviria: Orthornavirae), Including the Large Orders Bunyavirales and Mononegavirales
Archives of Virology https://doi.org/10.1007/s00705-020-04731-2 VIROLOGY DIVISION NEWS 2020 taxonomic update for phylum Negarnaviricota (Riboviria: Orthornavirae), including the large orders Bunyavirales and Mononegavirales Jens H. Kuhn1 · Scott Adkins2 · Daniela Alioto3 · Sergey V. Alkhovsky4 · Gaya K. Amarasinghe5 · Simon J. Anthony6,7 · Tatjana Avšič‑Županc8 · María A. Ayllón9,10 · Justin Bahl11 · Anne Balkema‑Buschmann12 · Matthew J. Ballinger13 · Tomáš Bartonička14 · Christopher Basler15 · Sina Bavari16 · Martin Beer17 · Dennis A. Bente18 · Éric Bergeron19 · Brian H. Bird20 · Carol Blair21 · Kim R. Blasdell22 · Steven B. Bradfute23 · Rachel Breyta24 · Thomas Briese25 · Paul A. Brown26 · Ursula J. Buchholz27 · Michael J. Buchmeier28 · Alexander Bukreyev18,29 · Felicity Burt30 · Nihal Buzkan31 · Charles H. Calisher32 · Mengji Cao33,34 · Inmaculada Casas35 · John Chamberlain36 · Kartik Chandran37 · Rémi N. Charrel38 · Biao Chen39 · Michela Chiumenti40 · Il‑Ryong Choi41 · J. Christopher S. Clegg42 · Ian Crozier43 · John V. da Graça44 · Elena Dal Bó45 · Alberto M. R. Dávila46 · Juan Carlos de la Torre47 · Xavier de Lamballerie38 · Rik L. de Swart48 · Patrick L. Di Bello49 · Nicholas Di Paola50 · Francesco Di Serio40 · Ralf G. Dietzgen51 · Michele Digiaro52 · Valerian V. Dolja53 · Olga Dolnik54 · Michael A. Drebot55 · Jan Felix Drexler56 · Ralf Dürrwald57 · Lucie Dufkova58 · William G. Dundon59 · W. Paul Duprex60 · John M. Dye50 · Andrew J. Easton61 · Hideki Ebihara62 · Toufc Elbeaino63 · Koray Ergünay64 · Jorlan Fernandes195 · Anthony R. Fooks65 · Pierre B. H. Formenty66 · Leonie F. Forth17 · Ron A. M. Fouchier48 · Juliana Freitas‑Astúa67 · Selma Gago‑Zachert68,69 · George Fú Gāo70 · María Laura García71 · Adolfo García‑Sastre72 · Aura R. Garrison50 · Aiah Gbakima73 · Tracey Goldstein74 · Jean‑Paul J. Gonzalez75,76 · Anthony Grifths77 · Martin H. Groschup12 · Stephan Günther78 · Alexandro Guterres195 · Roy A. -

X-Ray Structure of the Arenavirus Glycoprotein GP2 in Its Postfusion Hairpin Conformation
Corrections NEUROBIOLOGY Correction for “High-resolution structure of hair-cell tip links,” The authors note that Figure 3 appeared incorrectly. The by Bechara Kachar, Marianne Parakkal, Mauricio Kurc, Yi-dong corrected figure and its legend appear below. This error does not Zhao, and Peter G. Gillespie, which appeared in issue 24, affect the conclusions of the article. November 21, 2000, of Proc Natl Acad Sci USA (97:13336– 13341; 10.1073/pnas.97.24.13336). CORRECTIONS Fig. 3. Upper and lower attachments of the tip link. (A and B) Freeze-etch images of tip-link upper insertions in guinea pig cochlea (A) and (left to right) two from guinea pig cochlea, two from bullfrog sacculus, and two from guinea pig utriculus (B). Each example shows pronounced branching. (C and D) Freeze- etch images of the tip-link lower insertion in stereocilia from bullfrog sacculus (C) and guinea pig utriculus (D); multiple strands (arrows) arise from the stereociliary tip. (E) Freeze-fracture image of stereociliary tips from bullfrog sacculus; indentations at tips are indicated by arrows. (Scale bars: A = 100 nm, B = 25 nm; C–E = 100 nm.) www.pnas.org/cgi/doi/10.1073/pnas.1311228110 www.pnas.org PNAS | July 16, 2013 | vol. 110 | no. 29 | 12155–12156 Downloaded by guest on September 28, 2021 BIOCHEMISTRY BIOPHYSICS AND COMPUTATIONAL BIOLOGY, STATISTICS Correction for “X-ray structure of the arenavirus glycoprotein Correction for “Differential principal component analysis of GP2 in its postfusion hairpin conformation,” by Sébastien Igo- ChIP-seq,” by Hongkai Ji, Xia Li, Qian-fei Wang, and Yang net, Marie-Christine Vaney, Clemens Vonhrein, Gérard Bri- Ning, which appeared in issue 17, April 23, 2013, of Proc Natl cogne, Enrico A. -

MARBURG HEMORRHAGIC FEVER (Marburg HF)
MARBURG HEMORRHAGIC FEVER (Marburg HF) REPORTING INFORMATION • Class A: Report immediately via telephone the case or suspected case and/or a positive laboratory result to the local public health department where the patient resides. If patient residence is unknown, report immediately via telephone to the local public health department in which the reporting health care provider or laboratory is located. Local health departments should report immediately via telephone the case or suspected case and/or a positive laboratory result to the Ohio Department of Health (ODH). • Reporting Form(s) and/or Mechanism: o Immediately via telephone. o For local health departments, cases should also be entered into the Ohio Disease Reporting System (ODRS) within 24 hours of the initial telephone report to the ODH. • Key fields for ODRS reporting include: import status (whether the infection was travel-associated or Ohio-acquired), date of illness onset, and all the fields in the Epidemiology module. AGENT Marburg hemorrhagic fever is a rare, severe type of hemorrhagic fever which affects both humans and non-human primates. Caused by a genetically unique zoonotic RNA virus of the family Filoviridae, its recognition led to the creation of this virus family. The five species of Ebola virus are the only other known members of the family Filoviridae. Marburg virus was first recognized in 1967, when outbreaks of hemorrhagic fever occurred simultaneously in laboratories in Marburg and Frankfurt, Germany and in Belgrade, Yugoslavia (now Serbia). A total of 31 people became ill, including laboratory workers as well as several medical personnel and family members who had cared for them. -

Taxonomy of the Order Bunyavirales: Update 2019
Archives of Virology (2019) 164:1949–1965 https://doi.org/10.1007/s00705-019-04253-6 VIROLOGY DIVISION NEWS Taxonomy of the order Bunyavirales: update 2019 Abulikemu Abudurexiti1 · Scott Adkins2 · Daniela Alioto3 · Sergey V. Alkhovsky4 · Tatjana Avšič‑Županc5 · Matthew J. Ballinger6 · Dennis A. Bente7 · Martin Beer8 · Éric Bergeron9 · Carol D. Blair10 · Thomas Briese11 · Michael J. Buchmeier12 · Felicity J. Burt13 · Charles H. Calisher10 · Chénchén Cháng14 · Rémi N. Charrel15 · Il Ryong Choi16 · J. Christopher S. Clegg17 · Juan Carlos de la Torre18 · Xavier de Lamballerie15 · Fēi Dèng19 · Francesco Di Serio20 · Michele Digiaro21 · Michael A. Drebot22 · Xiaˇoméi Duàn14 · Hideki Ebihara23 · Toufc Elbeaino21 · Koray Ergünay24 · Charles F. Fulhorst7 · Aura R. Garrison25 · George Fú Gāo26 · Jean‑Paul J. Gonzalez27 · Martin H. Groschup28 · Stephan Günther29 · Anne‑Lise Haenni30 · Roy A. Hall31 · Jussi Hepojoki32,33 · Roger Hewson34 · Zhìhóng Hú19 · Holly R. Hughes35 · Miranda Gilda Jonson36 · Sandra Junglen37,38 · Boris Klempa39 · Jonas Klingström40 · Chūn Kòu14 · Lies Laenen41,42 · Amy J. Lambert35 · Stanley A. Langevin43 · Dan Liu44 · Igor S. Lukashevich45 · Tāo Luò1 · Chuánwèi Lüˇ 19 · Piet Maes41 · William Marciel de Souza46 · Marco Marklewitz37,38 · Giovanni P. Martelli47 · Keita Matsuno48,49 · Nicole Mielke‑Ehret50 · Maria Minutolo3 · Ali Mirazimi51 · Abulimiti Moming14 · Hans‑Peter Mühlbach50 · Rayapati Naidu52 · Beatriz Navarro20 · Márcio Roberto Teixeira Nunes53 · Gustavo Palacios25 · Anna Papa54 · Alex Pauvolid‑Corrêa55 · Janusz T. Pawęska56,57 · Jié Qiáo19 · Sheli R. Radoshitzky25 · Renato O. Resende58 · Víctor Romanowski59 · Amadou Alpha Sall60 · Maria S. Salvato61 · Takahide Sasaya62 · Shū Shěn19 · Xiǎohóng Shí63 · Yukio Shirako64 · Peter Simmonds65 · Manuela Sironi66 · Jin‑Won Song67 · Jessica R. Spengler9 · Mark D. Stenglein68 · Zhèngyuán Sū19 · Sùróng Sūn14 · Shuāng Táng19 · Massimo Turina69 · Bó Wáng19 · Chéng Wáng1 · Huálín Wáng19 · Jūn Wáng19 · Tàiyún Wèi70 · Anna E. -

Hong Kong's Bad Biodiversity
JULY 2001 NUMBER 23 Porcupine! Newsletter of the Department of Ecology & Biodiversity, The University of Hong Kong Hong Kong's Bad Biodiversity Aliens - also known as exotics or introduced species - are species which did not occur here naturally but have been brought to Hong Kong by humans, either deliberately, as crops, ornamentals, domestic animals or pets, or accidentally, as stowaways in ships and cargo planes, as seeds or eggs in soil, or attached to shoes or clothing. Most species which arrive in this way do not "escape" to form wild populations, and most of the species which have escaped are confined to places that are continually disturbed by humans. The weeds of cultivated areas, roadsides and urban wasteland are mostly exotics, as are the fish in concrete channels, and such conspicuous urban invertebrates as the "American" Cockroach and the Giant African Snail. Most of these urban exotics are doing no obvious harm and some contribute positively to the quality of city life. However, they also form the pool of species from which are drawn the small number of ecologically-damaging ones: the invasive species. Invasive species are aliens that invade natural or semi-natural communities. Out of the 150 or so well-established alien plant species in Hong Kong, less than a dozen have become invasive. The climbing American composite Mikania micrantha – Mile-a-Minute – is the most obvious example, but even this is confined to relatively disturbed sites. The "success" rate seems to be higher among birds and mammals, but the species which have done well in natural habitats are all from tropical East Asia and are apparently occupying niches vacated by the loss of the same or closely-related species when Hong Kong was deforested centuries ago. -

Taxonomy of the Order Bunyavirales: Second Update 2018
Archives of Virology (2019) 164:927–941 https://doi.org/10.1007/s00705-018-04127-3 VIROLOGY DIVISION NEWS Taxonomy of the order Bunyavirales: second update 2018 Piet Maes1 · Scott Adkins2 · Sergey V. Alkhovsky3 · Tatjana Avšič‑Županc4 · Matthew J. Ballinger5 · Dennis A. Bente6 · Martin Beer7 · Éric Bergeron8 · Carol D. Blair9 · Thomas Briese10 · Michael J. Buchmeier11 · Felicity J. Burt12,13 · Charles H. Calisher9 · Rémi N. Charrel14 · Il Ryong Choi15 · J. Christopher S. Clegg16 · Juan Carlos de la Torre17 · Xavier de Lamballerie14 · Joseph L. DeRisi18,19,20 · Michele Digiaro21 · Mike Drebot22 · Hideki Ebihara23 · Toufc Elbeaino21 · Koray Ergünay24 · Charles F. Fulhorst6 · Aura R. Garrison25 · George Fú Gāo26 · Jean‑Paul J. Gonzalez27 · Martin H. Groschup28,29 · Stephan Günther30 · Anne‑Lise Haenni31 · Roy A. Hall32 · Roger Hewson33 · Holly R. Hughes34 · Rakesh K. Jain35 · Miranda Gilda Jonson36 · Sandra Junglen37,38 · Boris Klempa37,39 · Jonas Klingström40 · Richard Kormelink41 · Amy J. Lambert34 · Stanley A. Langevin42 · Igor S. Lukashevich43 · Marco Marklewitz37,38 · Giovanni P. Martelli44 · Nicole Mielke‑Ehret45 · Ali Mirazimi46 · Hans‑Peter Mühlbach45 · Rayapati Naidu47 · Márcio Roberto Teixeira Nunes48 · Gustavo Palacios25 · Anna Papa49 · Janusz T. Pawęska50,51 · Clarence J. Peters6 · Alexander Plyusnin52 · Sheli R. Radoshitzky25 · Renato O. Resende53 · Víctor Romanowski54 · Amadou Alpha Sall55 · Maria S. Salvato56 · Takahide Sasaya57 · Connie Schmaljohn25 · Xiǎohóng Shí58 · Yukio Shirako59 · Peter Simmonds60 · Manuela Sironi61 · -

Past, Present, and Future of Arenavirus Taxonomy
Arch Virol DOI 10.1007/s00705-015-2418-y VIROLOGY DIVISION NEWS Past, present, and future of arenavirus taxonomy Sheli R. Radoshitzky1 · Yīmíng Bào2 · Michael J. Buchmeier3 · Rémi N. Charrel4,18 · Anna N. Clawson5 · Christopher S. Clegg6 · Joseph L. DeRisi7,8,9 · Sébastien Emonet10 · Jean-Paul Gonzalez11 · Jens H. Kuhn5 · Igor S. Lukashevich12 · Clarence J. Peters13 · Victor Romanowski14 · Maria S. Salvato15 · Mark D. Stenglein16 · Juan Carlos de la Torre17 © Springer-Verlag Wien 2015 Abstract Until recently, members of the monogeneric Arenaviridae to accommodate reptilian arenaviruses and family Arenaviridae (arenaviruses) have been known to other recently discovered mammalian arenaviruses and to infect only muroid rodents and, in one case, possibly improve compliance with the Rules of the International phyllostomid bats. The paradigm of arenaviruses exclu- Code of Virus Classification and Nomenclature (ICVCN). sively infecting small mammals shifted dramatically when PAirwise Sequence Comparison (PASC) of arenavirus several groups independently published the detection and genomes and NP amino acid pairwise distances support the isolation of a divergent group of arenaviruses in captive modification of the present classification. As a result, the alethinophidian snakes. Preliminary phylogenetic analyses current genus Arenavirus is replaced by two genera, suggest that these reptilian arenaviruses constitute a sister Mammarenavirus and Reptarenavirus, which are estab- clade to mammalian arenaviruses. Here, the members of lished to accommodate mammalian and reptilian the International Committee on Taxonomy of Viruses arenaviruses, respectively, in the same family. The current (ICTV) Arenaviridae Study Group, together with other species landscape among mammalian arenaviruses is experts, outline the taxonomic reorganization of the family upheld, with two new species added for Lunk and Merino Walk viruses and minor corrections to the spelling of some names. -

Taxonomy of the Family Arenaviridae and the Order Bunyavirales: Update 2018
Archives of Virology https://doi.org/10.1007/s00705-018-3843-5 VIROLOGY DIVISION NEWS Taxonomy of the family Arenaviridae and the order Bunyavirales: update 2018 Piet Maes1 · Sergey V. Alkhovsky2 · Yīmíng Bào3 · Martin Beer4 · Monica Birkhead5 · Thomas Briese6 · Michael J. Buchmeier7 · Charles H. Calisher8 · Rémi N. Charrel9 · Il Ryong Choi10 · Christopher S. Clegg11 · Juan Carlos de la Torre12 · Eric Delwart13,14 · Joseph L. DeRisi15 · Patrick L. Di Bello16 · Francesco Di Serio17 · Michele Digiaro18 · Valerian V. Dolja19 · Christian Drosten20,21,22 · Tobiasz Z. Druciarek16 · Jiang Du23 · Hideki Ebihara24 · Toufc Elbeaino18 · Rose C. Gergerich16 · Amethyst N. Gillis25 · Jean‑Paul J. Gonzalez26 · Anne‑Lise Haenni27 · Jussi Hepojoki28,29 · Udo Hetzel29,30 · Thiện Hồ16 · Ní Hóng31 · Rakesh K. Jain32 · Petrus Jansen van Vuren5,33 · Qi Jin34,35 · Miranda Gilda Jonson36 · Sandra Junglen20,22 · Karen E. Keller37 · Alan Kemp5 · Anja Kipar29,30 · Nikola O. Kondov13 · Eugene V. Koonin38 · Richard Kormelink39 · Yegor Korzyukov28 · Mart Krupovic40 · Amy J. Lambert41 · Alma G. Laney42 · Matthew LeBreton43 · Igor S. Lukashevich44 · Marco Marklewitz20,22 · Wanda Markotter5,33 · Giovanni P. Martelli45 · Robert R. Martin37 · Nicole Mielke‑Ehret46 · Hans‑Peter Mühlbach46 · Beatriz Navarro17 · Terry Fei Fan Ng14 · Márcio Roberto Teixeira Nunes47,48 · Gustavo Palacios49 · Janusz T. Pawęska5,33 · Clarence J. Peters50 · Alexander Plyusnin28 · Sheli R. Radoshitzky49 · Víctor Romanowski51 · Pertteli Salmenperä28,52 · Maria S. Salvato53 · Hélène Sanfaçon54 · Takahide Sasaya55 · Connie Schmaljohn49 · Bradley S. Schneider25 · Yukio Shirako56 · Stuart Siddell57 · Tarja A. Sironen28 · Mark D. Stenglein58 · Nadia Storm5 · Harikishan Sudini59 · Robert B. Tesh48 · Ioannis E. Tzanetakis16 · Mangala Uppala59 · Olli Vapalahti28,30,60 · Nikos Vasilakis48 · Peter J. Walker61 · Guópíng Wáng31 · Lìpíng Wáng31 · Yànxiăng Wáng31 · Tàiyún Wèi62 · Michael R. -

Genomic Diversity and Spatiotemporal Distributions of Lassa Virus Outbreaks in Nigeria
Genomic Diversity and Spatiotemporal Distributions of Lassa Virus Outbreaks in Nigeria Abdulwahid Abaukaka Yahaya Tehran University of Medical Sciences Yazdan Asgari ( [email protected] ) Tehran University of Medical Sciences https://orcid.org/0000-0001-6993-6956 Research article Keywords: Genomic, Lassa Virus, Phylogeography, Spatiotemporal and Nigeria Posted Date: October 28th, 2019 DOI: https://doi.org/10.21203/rs.2.16266/v2 License: This work is licensed under a Creative Commons Attribution 4.0 International License. Read Full License Page 1/20 Abstract Abstract Background Lassa virus (LASV) is a single-negative strand RNA Arenavirus (genus Mammarenavirus), oriented in both negative and positive senses. Due to the increase in the fatality rate of deadly disease LASV caused (Lassa fever), widespread LASV in Nigeria has been a subject of interest. Following the upsurge of LASV endemicity in 2012, another marked incidence recorded in Nigeria, 2018, with 394 conrmed cases in 19 states, and an estimated 25% cases led to death. This study aimed at acquiring the genetic variation of LASV ancestral evolution with the evolvement of new strains in different lineage and its geographical distributions within a specic time of outbreaks through Bayesian inference, using genomic sequence across affected states in Nigeria. Results From the result, we were able to establish the relationship of Lassa mamarenavirus and other arenaviruses by classifying them into distinct monophyletic groups, i.e., the old world arenaviruses, new world arenaviruses, and Reptarenaviruses. Corresponding promoter sites for genetic expression of the viral genome were analyzed based on Transcription Starting Site (TSS), the S_Segment (MK291249.1) is about 2917–2947 bp and L_Segment (MH157036.1), is about1863–1894 bp long. -
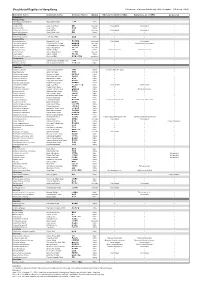
Checklist of Reptiles in Hong Kong © Programme of Ecology & Biodiversity, HKU (Last Update: 10 September 2012)
Checklist of Reptiles in Hong Kong © Programme of Ecology & Biodiversity, HKU (Last Update: 10 September 2012) Scientific name Common name Chinese Name Status Ades & Kendrick (2004) Karsen et al. (1998) Uetz et al. Testudines Platysternidae Platysternon megacephalum Big-headed Terrapin 大頭龜 Native Cheloniidae Caretta caretta Loggerhead Turtle 蠵龜 Uncertain Not included Not included Chelonia mydas Green Turtle 緣海龜 Native Eretmochelys imbricata Hawksbill Turtle 玳瑁 Uncertain Not included Not included Lepidochelys olivacea Pacific Ridley Turtle 麗龜 Native Dermochelyidae Dermochelys coriacea Leatherback Turtle 稜皮龜 Native Geoemydidae Cuora amboinensis Malayan Box Turtle 馬來閉殼龜 Introduced Not included Not included Cuora flavomarginata Yellow-lined Box Terrapin 黃緣閉殼龜 Uncertain Cistoclemmys flavomarginata Cuora trifasciata Three-banded Box Terrapin 三線閉殼龜 Native Mauremys mutica Chinese Pond Turtle 黃喉水龜 Uncertain Mauremys reevesii Reeves' Terrapin 烏龜 Native Chinemys reevesii Chinemys reevesii Ocadia sinensis Chinese Striped Turtle 中華花龜 Uncertain Sacalia bealei Beale's Terrapin 眼斑水龜 Native Trachemys scripta elegans Red-eared Slider 巴西龜 / 紅耳龜 Introduced Trionychidae Palea steindachneri Wattle-necked Soft-shelled Turtle 山瑞鱉 Uncertain Pelodiscus sinensis Chinese Soft-shelled Turtle 中華鱉 / 水魚 Native Squamata - Serpentes Colubridae Achalinus rufescens Rufous Burrowing Snake 棕脊蛇 Native Achalinus refescens (typo) Ahaetulla prasina Jade Vine Snake 綠瘦蛇 Uncertain Amphiesma atemporale Mountain Keelback 無顳鱗游蛇 Native -

List of Reptile Species in Hong Kong
List of Reptile Species in Hong Kong Family No. of Species Common Name Scientific Name Order TESTUDOFORMES Cheloniidae 4 Loggerhead Turtle Caretta caretta Green Turtle Chelonia mydas Hawksbill Turtle Eretmochelys imbricata Olive Ridley Turtle Lepidochelys olivacea Dermochelyidae 1 Leatherback Turtle Dermochelys coriacea Emydidae 1 Red-eared Slider * Trachemys scripta elegans Geoemydidae 3 Three-banded Box Turtle Cuora trifasciata Reeves' Turtle Mauremys reevesii Beale's Turtle Sacalia bealei Platysternidae 1 Big-headed Turtle Platysternon megacephalum Trionychidae 1 Chinese Soft-shelled Turtle Pelodiscus sinensis Order SQUAMATA Suborder LACERTILIA Agamidae 1 Changeable Lizard Calotes versicolor Lacertidae 1 Grass Lizard Takydromus sexlineatus ocellatus Scincidae 11 Chinese Forest Skink Ateuchosaurus chinensis Long-tailed Skink Eutropis longicaudata Chinese Skink Plestiodon chinensis chinensis Five-striped Blue-tailed Skink Plestiodon elegans Blue-tailed Skink Plestiodon quadrilineatus Vietnamese Five-lined Skink Plestiodon tamdaoensis Slender Forest Skink Scincella modesta Reeve's Smooth Skink Scincella reevesii Brown Forest Skink Sphenomorphus incognitus Indian Forest Skink Sphenomorphus indicus Chinese Waterside Skink Tropidophorus sinicus Varanidae 1 Common Water Monitor Varanus salvator Dibamidae 1 Bogadek's Burrowing Lizard Dibamus bogadeki Gekkonidae 8 Four-clawed Gecko Gehyra mutilata Chinese Gecko Gekko chinensis Tokay Gecko Gekko gecko Bowring's Gecko Hemidactylus bowringii Brook's Gecko* Hemidactylus brookii House Gecko* Hemidactylus