Illinois Natural History Survey
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Vocalization Behavior of the Endangered Bahama Oriole (Icterus Northropi): Ontogenetic, Sexual, Temporal, Duetting Pair, and Geographic Variation Valerie A
Loma Linda University TheScholarsRepository@LLU: Digital Archive of Research, Scholarship & Creative Works Loma Linda University Electronic Theses, Dissertations & Projects 3-1-2011 Vocalization Behavior of the Endangered Bahama Oriole (Icterus northropi): Ontogenetic, Sexual, Temporal, Duetting Pair, and Geographic Variation Valerie A. Lee Loma Linda University Follow this and additional works at: http://scholarsrepository.llu.edu/etd Part of the Biology Commons Recommended Citation Lee, Valerie A., "Vocalization Behavior of the Endangered Bahama Oriole (Icterus northropi): Ontogenetic, Sexual, Temporal, Duetting Pair, and Geographic Variation" (2011). Loma Linda University Electronic Theses, Dissertations & Projects. 37. http://scholarsrepository.llu.edu/etd/37 This Thesis is brought to you for free and open access by TheScholarsRepository@LLU: Digital Archive of Research, Scholarship & Creative Works. It has been accepted for inclusion in Loma Linda University Electronic Theses, Dissertations & Projects by an authorized administrator of TheScholarsRepository@LLU: Digital Archive of Research, Scholarship & Creative Works. For more information, please contact [email protected]. LOMA LINDA UNIVERSITY School of Science and Technology in conjunction with the Faculty of Graduate Studies ____________________ Vocalization Behavior of the Endangered Bahama Oriole (Icterus northropi): Ontogenetic, Sexual, Temporal, Duetting Pair, and Geographic Variation by Valerie A. Lee ____________________ A Thesis submitted in partial satisfaction of the requirements for the degree of Master of Science in Biology ____________________ March 2011 © 2011 Valerie A. Lee All Rights Reserved Each person whose signature appears below certifies that this thesis in his/her opinion is adequate, in scope and quality, as a thesis for the degree Master of Science. , Chairperson William K. Hayes, Professor of Biology Stephen G. -

Interactions Between Migrating Birds and Offshore Oil and Gas Platforms in the Northern Gulf of Mexico
OCS Study MMS 2005-009 Coastal Marine Institute Interactions Between Migrating Birds and Offshore Oil and Gas Platforms in the Northern Gulf of Mexico Final Report U.S. Department of the Interior Cooperative Agreement Minerals Management Service Coastal Marine Institute Gulf of Mexico OCS Region Louisiana State University OCS Study MMS 2005-009 Coastal Marine Institute Interactions Between Migrating Birds and Offshore Oil and Gas Platforms in the Northern Gulf of Mexico Final Report Editor Robert W. Russell February 2005 Prepared under MMS Contract 1435-01-99-CA-30951-16808 by The Migration Over the Gulf Project School of the Coast and Environment Louisiana State University Baton Rouge, Louisiana 70803 Published by U.S. Department of the Interior Cooperative Agreement Minerals Management Service Coastal Marine Institute Gulf of Mexico OCS Region Louisiana State University DISCLAIMER This report was prepared under contract between the Minerals Management Service (MMS) and Louisiana State University. This report has been technically reviewed by the MMS, and it has been approved for publication. Approval does not signify that the contents necessarily reflect the views and policies of the MMS, nor does mention of trade names or commercial products constitute endorsement or recommendation for use. It is, however, exempt from review and compliance with the MMS editorial standards. REPORT AVAILABILITY Extra copies of this report may be obtained from the Public Information Office at the following address: U.S. Department of the Interior Minerals Management Service Gulf of Mexico OCS Region Public Information Office (MS 5034) 1201 Elmwood Park Boulevard New Orleans, LA 70123-2394 Telephone: (504) 736-2519 or (800) 200-GULF CITATION Suggested citation: Russell, R.W. -

Song and Plumage Evolution in the New World Orioles (Icterus) Show Similar Lability and Convergence in Patterns
ORIGINAL ARTICLE doi:10.1111/j.1558-5646.2007.00082.x SONG AND PLUMAGE EVOLUTION IN THE NEW WORLD ORIOLES (ICTERUS) SHOW SIMILAR LABILITY AND CONVERGENCE IN PATTERNS J. Jordan Price,1,2 Nicholas R. Friedman,1,3 and Kevin E. Omland4,5 1Department of Biology, St. Mary’s College of Maryland, St. Mary’s City, Maryland 20686 2E-mail: [email protected] 3E-mail: [email protected] 4Department of Biological Sciences, University of Maryland, Baltimore County, Baltimore, Maryland 21250 5E-mail: [email protected] Received August 28, 2006 Accepted November 23, 2006 Both song and color patterns in birds are thought to evolve rapidly and exhibit high levels of homoplasy, yet few previous studies have compared the evolution of these traits systematically using the same taxa. Here we reconstruct the evolution of song in the New World orioles (Icterus) and compare patterns of vocal evolution to previously reconstructed patterns of change in plumage evolution in this clade. Individual vocal characters exhibit high levels of homoplasy, reflected in a low overall consistency index (CI = 0.27) and retention index (RI = 0.35). Levels of lability in song are comparable to those found for oriole plumage patterns using the same taxa (CI = 0.31, RI = 0.63), but are strikingly dissimilar to the conservative patterns of change seen in the songs of oropendolas (Psarocolius, Ocyalus;CI= 0.82, RI = 0.87), a group closely related to the orioles. Oriole song is also similar to oriole plumage in exhibiting repeated convergence in overall patterns, with some distantly related taxa sounding remarkably similar. -

Notes on the Ornithology of Southern Texas
University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Papers in Ornithology Papers in the Biological Sciences 1878 Notes on the Ornithology of Southern Texas James C. Merrill U.S. Army Follow this and additional works at: https://digitalcommons.unl.edu/biosciornithology Part of the Ornithology Commons Merrill, James C., "Notes on the Ornithology of Southern Texas" (1878). Papers in Ornithology. 55. https://digitalcommons.unl.edu/biosciornithology/55 This Article is brought to you for free and open access by the Papers in the Biological Sciences at DigitalCommons@University of Nebraska - Lincoln. It has been accepted for inclusion in Papers in Ornithology by an authorized administrator of DigitalCommons@University of Nebraska - Lincoln. o ES O.T THE OF SOUTHER UM. n A I T OF BIRD~ OB1IER\ ED I THE VICI IT OF FORT BROW/, TIE AI FRO:\I ,1876,'IOJ , 1 7 . By JAMES C. MERRILL, G OFF1CE. NOTES . ON THE ORNITHOLOGY OF SOUTHERN TEXAS, BEING A LIST OF BIRDS OBSERVED IN THE VICINITY OF FORT BROWN, TEXAS, FROM FEBRUARY, 1876, TO JUN~, 1878'. ,By JAMES C. MERRILL, ASSISTANT SURGEON U. S. ARMY. WASHINGTON: GOVERNMENT PRINTING OFFICE. 1878. 118 PIWCEEDINGS OF UNITED STATES NATION.\L MUSEU~1. NOTES ON THE ORNITHOLOGY OF SOUTHERN TEXAS, BEING A LIST OF BIRD~ OBSERVED IX TilE V.lrUIYITY OF FORT BROWN, TEXAS, FROM FEBRUARY, 1816, TO JUNE. 1818. By .JAMES (J. lJIERRILL, Assistant Surgeon U. S. Army. The post of' Fort Brown, Texas, in the immediate vicinity of which most of the following observations were maoe, is 'at the extreme southern point of the State, in latitude 250 53' IG", longitude 97 0 13'. -

Pre-Lesson Plan
Pre-Lesson Plan Prior to taking part in the Winged Migration program at Tommy Thompson Park it is recommended that you complete the following lessons to familiarize your students with some of the birds they might see and some of the concepts they will learn during their field trip. The lessons can easily be integrated into your Science, Language Arts, Social Studies and Physical Education programs. Part 1: Amazing Birds As a class, read the provided “Wanted” posters. The posters depict a very small sampling of some of the amazing feats and features of birds. To complement these readings, display the following websites so that students can see some of these birds “up close.” Common Loon http://www.schollphoto.com/gallery/thumbnails.php?album=1 Black-Capped Chickadee http://sdakotabirds.com/species_photos/black_capped_chickadee.htm Ruby-Throated Hummingbird http://www.surfbirds.com/cgi-bin/gallery/search2.cgi?species=Ruby- throated%20Hummingbird Downy Woodpecker http://www.pbase.com/billko/downy_woodpecker Great Horned Owl www.owling.com/Great_Horned.htm When you visit Tommy Thompson Park, you may see chickadees, hummingbirds, and woodpeckers. These birds all breed in southern Ontario. However, you probably will not see a Great Horned Owl, because this specific bird is usually flying around at night. Below is a list of some other birds students might see when they visit Tommy Thompson Park. Have them chose one bird each and write a “Wanted” poster for it, focusing on a cool fact about that bird. Some web sites that will help them get started -

RETURN of NORTHERN MIGRANTS to TROPICAL WINTER QUARTERS and BANDED BIRDS RECOVERED in the UNITED STATES by Walter P
RETURN OF NORTHERN MIGRANTS TO TROPICAL WINTER QUARTERS AND BANDED BIRDS RECOVERED IN THE UNITED STATES By Walter P. Nickell There are many records of long-distance migrants returning one or more years to the same nesting locality and even to identical nest sites in the north. Further, there are within the last fifty years, numerous records in the literature of farther-north migrants such as the Tree Sparrow (Spizella arborea) and Slate-colored Junco (Junco hyemalis), which have returned to the same winter quarters at lower latitudes but not into the subtropics or tropics. Records of the return of northern migrants to identical tropical winter quarters are not common, probably because only within recent years have researchers in the tropics done extensive banding through the use of mist nets, to continue banding in the same net sets in successive years. As early as 1915 Baldwin, running a trap ping station at Thomasville, Georgia, began to record such species as the White-throated Sparrow (Zonotrichia albicollis), White- crowned Sparrow (Zonotrichia leucophrys), Chipping Sparrow (Spizella passerina), Song Sparrow (Melospiza melodia), Hermit Thrush (Hylocichla guttata), and Myrtle Warbler (Dendroica coronata) (all undoubted migrants) as returning for several suc cessive years to the same trapping station. Apparently the first proof that migrants to the tropics often return to the same winter grounds was obtained by the late J. Van Tyne and A. Ledyard Smith (p. 110). Van Tyne banded 99 Indigo Buntings (Passerina cyanea) at Uaxactun, Guatemala during April 1931. In April 1932, at the same spot, Smith trapped six of the Buntings banded the year before. -

Victory for the Quads!
DECEMBER 2019 MONTHLY MEETING Monday, December 9th The Sarasota Audubon Society Nature Center Open daily 9:00am-1:00pm Click Through This Issue! :: MONTHLY MEETING :: SAS 2019-20 SCHEDULE :: SAS ACTIVITIES THE MYSTERY AND MAGIC OF :: CLASSES AND WORKSHOPS SARASTOA'S SWALLOW-TAILED :: FOR THE BIRDS :: WRITE THE EDITOR KITES - PART TWO :: RENEW MEMBERSHIP :: CONSERVATION Margi Haas, Citizen Scientist and Kite Fanatic will :: VOLUNTEER continue her discussions about Swallow-tailed Kites. :: WHAT'S NEW AT THE NC :: SAS GIFT SHOP Social Hour: 6:30pm :: SPOONBILL CLUB Business Meeting: 7:00pm :: SAS COMMEMORATIVE BRICKS Presentation: 7:30pm :: SUBMIT AN ARTICLE :: SAS WEB SITE First Congregational Church Fellowship Hall 2031 S. Euclid, Sarasota VICTORY FOR THE QUADS! ******** CLICK -> MONTHLY MEETING PROGRAMS ******* Pre-Monthly Meeting Dinner All Members Welcome! Gecko's Restaurant 1900 Hillview Ave 5:00pm Contact Karen Jensen ([email protected]) 2019-2020 ACTIVITY SCHEDULE CLICK FOR COMPLETE SCHEDULE SCHEDULE An Astounding Outcome! On November 6, 2019, the Board of County Commissioners of Sarasota County approved a conservation easement for 3 of the 4 Quad parcels SAS ACTIVITIES at the corners of Palmer Blvd and Apex/Coburn Roads in the Celery CLICK --> COMPLETE SCHEDULE Fields. We were thrilled and amazed. Thanks to Fresh Start for all of their efforts towards this combined and important step for the environment. GUIDED WALKS In partnership with the Conservation Foundation of the Gulf Coast, Our SAS guided walks are led by some of our most Sarasota Audubon will work to, in effect, extend the Celery Fields knowledgeable and experienced birders. Come along another 30+ acres, creating a buffer zone for birds and wildlife to thrive, with us! and setting aside areas for low impact public use. -

MICHIGAN BIRDS and Natural History
MICHIGAN BIRDS and Natural History November-December 2010 Volume 17 Number 5 A publication of Michigan Audubon MICHIGAN BIRDS and Natural History November - December 2010 Volume 17 Number 5 Michigan Birds and Natural History is a publication of Michigan Audubon and is published five times a year. Its mission is to provide a forum for the publication of research on the natural history of Michigan with an emphasis on birds. It is also the home of the Michigan Bird Survey, Michigan Butterfly Survey, Michigan Christmas Bird Counts, North American Migration Count, and the proceedings of the Michigan Bird Records Committee. Subscriptions are $25.00 per year in the United States and $30.00 elsewhere. They cover the current calendar year. Subscriptions can be obtained by sending a check along with your name and address to: Michigan Audubon/MBNH 6011 W. St. Joseph Hwy., Suite 403 Lansing, Michigan 48917 MBNH Committee Managing Editor ............................................................................Jon Wuepper Layout Editor .................................................................................... Don Tinson II Survey Editor .............................................................................. Adam M. Byrne Photo Editor ............................................................................... Allen T. Chartier Copy Editor ............................................................................................ Bill Rapai Photo Quiz Editor ............................................................................. -
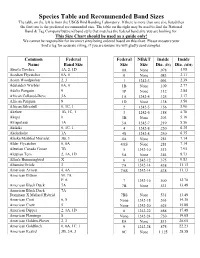
Species Table and Recommended Band Sizes the Table on the Left Is from the USGS Bird Banding Laboratory
Species Table and Recommended Band Sizes The table on the left is from the USGS Bird Banding Laboratory. If there is more than one size listed then the first one is the preferred recommended size. The table on the right may be used to find the National Band & Tag Company butt-end band style that matches the federal band size you are looking for. This Size Chart should be used as a guide only! We cannot be responsible for incorrect sizes being ordered based on this chart. Please measure your bird’s leg for accurate sizing, if you are unsure we will gladly send samples. Common Federal Federal NB&T Inside Inside Name Band Size Size Size Dia. (IN) Dia. (MM) Abert's Towhee 1A, 2, 1D 0A None .078 1.98 Acadian Flycatcher 0A, 0 0 None .083 2.11 Acorn Woodpecker 2, 3 1 1242-3 .094 2.39 Adelaide's Warbler 0A, 0 1B None .109 2.77 Adelie Penguin 9 1P None .112 2.84 African Collared-Dove 3A 1A 1242-4 .125 3.17 African Penguin 9 1D None .138 3.50 African Silverbill 0, 1C, 1 2 1242-5 .156 3.96 Akekee 1B, 1C, 1 3 1242-6 .188 4.78 Akepa 0 3B None .203 5.16 Akiapolaau 1A 3A 1242-7 .219 5.56 Akikiki 0, 1C, 1 4 1242-8 .250 6.35 Akohekohe 1A 4S 1242-8 .250 6.35 Alaska Marbled Murrelet 3B, 3 4A None .281 7.14 Alder Flycatcher 0, 0A 4AS None .281 7.14 Aleutian Canada Goose 7B 5 1242-10 .313 7.95 Aleutian Tern 2, 1A, 1D 5A None .344 8.73 Allen's Hummingbird X 6 1242-12 .375 9.53 Altamira Oriole 3 7A 1242-14 .438 11.13 American Avocet 4, 4A 7AS 1242-14 .438 11.13 American Bittern M: 7A F: 6 7 1242-16 .500 12.70 American Black Duck 7A 7B None .531 13.49 American -

Learn About Texas Birds Activity Book
Learn about . A Learning and Activity Book Color your own guide to the birds that wing their way across the plains, hills, forests, deserts and mountains of Texas. Text Mark W. Lockwood Conservation Biologist, Natural Resource Program Editorial Direction Georg Zappler Art Director Elena T. Ivy Educational Consultants Juliann Pool Beverly Morrell © 1997 Texas Parks and Wildlife 4200 Smith School Road Austin, Texas 78744 PWD BK P4000-038 10/97 All rights reserved. No part of this work covered by the copyright hereon may be reproduced or used in any form or by any means – graphic, electronic, or mechanical, including photocopying, recording, taping, or information storage and retrieval systems – without written permission of the publisher. Another "Learn about Texas" publication from TEXAS PARKS AND WILDLIFE PRESS ISBN- 1-885696-17-5 Key to the Cover 4 8 1 2 5 9 3 6 7 14 16 10 13 20 19 15 11 12 17 18 19 21 24 23 20 22 26 28 31 25 29 27 30 ©TPWPress 1997 1 Great Kiskadee 16 Blue Jay 2 Carolina Wren 17 Pyrrhuloxia 3 Carolina Chickadee 18 Pyrrhuloxia 4 Altamira Oriole 19 Northern Cardinal 5 Black-capped Vireo 20 Ovenbird 6 Black-capped Vireo 21 Brown Thrasher 7Tufted Titmouse 22 Belted Kingfisher 8 Painted Bunting 23 Belted Kingfisher 9 Indigo Bunting 24 Scissor-tailed Flycatcher 10 Green Jay 25 Wood Thrush 11 Green Kingfisher 26 Ruddy Turnstone 12 Green Kingfisher 27 Long-billed Thrasher 13 Vermillion Flycatcher 28 Killdeer 14 Vermillion Flycatcher 29 Olive Sparrow 15 Blue Jay 30 Olive Sparrow 31 Great Horned Owl =female =male Texas Birds More kinds of birds have been found in Texas than any other state in the United States: just over 600 species. -

27 Subpart C—Addresses
U.S. Fish and Wildlife Serv., Interior § 10.22 Quiscalus quiscula, Common Grackle fax Drive, Room 700, Arlington, VA Quiscalus niger, Greater Antillean Grackle 22203. Address mail for the following Molothrus bonariensis, Shiny Cowbird permits to the Regional Director. In Molothrus aeneus, Bronzed Cowbird Molothrus ater, Brown-headed Cowbird the address include one of the fol- Icterus dominicensis, Black-cowled Oriole lowing: for import/export licenses and Icterus wagleri, Black-vented Oriole exception to designated port permits Icterus spurius, Orchard Oriole (Attention: Import/export license); for Icterus cucullatus, Hooded Oriole native endangered and threatened spe- Icterus pustulatus, Streak-backed Oriole cies (Attention: Endangered/threatened Icterus gularis, Altamira Oriole species permit); and for migratory Icterus graduacauda, Audubon’s Oriole Icterus galbula, Northern Oriole birds and eagles (Attention: Migratory Icterus parisorum, Scott’s Oriole bird permit office). You can find ad- FAMILY FRINGILLIDAE dresses for regional offices at 50 CFR SUBFAMILY FRINGILLINAE 2.2. Fringilla montifringilla, Brambling [55 FR 48851, Nov. 23, 1990, as amended at 63 SUBFAMILY CARDUELINAE FR 52633, Oct. 1, 1998] Leucosticte arctoa, Rosy Finch Pinicola enucleator, Pine Grosbeak § 10.22 Law enforcement offices. Carpodacus erythrinus, Common Rosefinch Service law enforcement offices and Carpodacus purpureus, Purple Finch their areas of responsibility follow. Carpodacus cassinii, Cassin’s Finch Carpodacus mexicanus, House Finch Mail should be addressed: ‘‘Assistant Loxia curvirostra, Red Crossbill Regional Director, Division of Law En- Loxia leucoptera, White-winged Crossbill forcement, U.S. Fish and Wildlife Serv- Carduelis flammea, Common Redpoll ice, (appropriate address below)’’: Carduelis hornemanni, Hoary Redpoll Carduelis pinus, Pine Siskin AREAS OF RESPONSIBILITY AND OFFICE Carduelis psaltria, Lesser Goldfinch ADDRESSES Carduelis lawrencei, Lawrence’s Goldfinch California, Hawaii, Idaho. -

Observations on the Orchard Oriole in Lower Mississippi Delta
12• DENNIS,Observations onthe Orchard Oriole JalluaryBird-Banding 6. The oldest Oven-bird recorded during the study was a male which reached the age of at least seven years, one year more than the oldest known previous record. Its age equals that of each of four other warblers, a Yellow Warbler, Myrtle Warbler, Pine Warbler, and a Maryland Yellowthroat, which may indicate that the longevity in these species and perhaps in other warblers is similar to that of the Oven-bird. REFERENCES BURKITT, J. P., 1926. A study of the Robin by means of marked birds. Brit. Birds, 20: 91-101. CooK•:, .Nl.aYT. 1937. Some longevity recordsof wild birds. Bird-Banding. 8: 52-65. F.•RN•;R, I)o•'^•) S. 1945. Age groups and longevity in the American Robin. Wilson B,ll., 57: 56-74. GI•, Guov'•'R•¾. 1944. Old age-records of banded birds--9th list. Ebba News, 7, no. 12. ---- 1945. Old age-recordsof banded birds -- 10th list. Ebba News, 8. no. 2. ---- 1946. Old age-recordsof banded birds -- Final list. Ebba News, 9, no. 3. ------ 1947. Recovery page. Ebba News, 10, no. 2. HANN, HARRY W. 1937. Life history of the Oven-bird in southern Michigan. Wilson B•dl., 49: 145-237. -- 1940. Polyandry in the Oven-bird. Wilson Bull., 52: 69-72. -- 1941. The Cowbird at the nest. Wilson Bull., 53: 211-221. LAC•, D.avli). 1943a. The age of the Blackbird. Brit. Birds, 36: 166-175. ---- 1943b. The age of some more British birds. Brit. Birds, 36: 193-197; 214-221. ---- 1943c. The life of the Robin.