Journal.Pone.0146782
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Parish Share Report
PARISH SHARE PAYMENTS For period ended 30th September 2019 SUMMARY OF PARISH SHARE PAYMENTS BY DEANERIES Dean Amount % Deanery Share Received for 2019 % Deanery Share % No Outstanding 2018 2019 to period end 2018 Received for 2018 received £ £ £ £ £ Norwich Archdeaconry 06 Norwich East 23,500 4.41 557,186 354,184 63.57 532,380 322,654 60.61 04 Norwich North 47,317 9.36 508,577 333,671 65.61 505,697 335,854 66.41 05 Norwich South 28,950 7.21 409,212 267,621 65.40 401,270 276,984 69.03 Norfolk Archdeaconry 01 Blofield 37,303 11.04 327,284 212,276 64.86 338,033 227,711 67.36 11 Depwade 46,736 16.20 280,831 137,847 49.09 288,484 155,218 53.80 02 Great Yarmouth 44,786 9.37 467,972 283,804 60.65 478,063 278,114 58.18 13 Humbleyard 47,747 11.00 437,949 192,301 43.91 433,952 205,085 47.26 14 Loddon 62,404 19.34 335,571 165,520 49.32 322,731 174,229 53.99 15 Lothingland 21,237 3.90 562,194 381,997 67.95 545,102 401,890 73.73 16 Redenhall 55,930 17.17 339,813 183,032 53.86 325,740 187,989 57.71 09 St Benet 36,663 9.24 380,642 229,484 60.29 396,955 243,433 61.33 17 Thetford & Rockland 31,271 10.39 314,266 182,806 58.17 300,933 192,966 64.12 Lynn Archdeaconry 18 Breckland 45,799 11.97 397,811 233,505 58.70 382,462 239,714 62.68 20 Burnham & Walsingham 63,028 15.65 396,393 241,163 60.84 402,850 256,123 63.58 12 Dereham in Mitford 43,605 12.03 353,955 223,631 63.18 362,376 208,125 57.43 21 Heacham & Rising 24,243 6.74 377,375 245,242 64.99 359,790 242,156 67.30 22 Holt 28,275 8.55 327,646 207,089 63.21 330,766 214,952 64.99 23 Lynn 10,805 3.30 330,152 196,022 59.37 326,964 187,510 57.35 07 Repps 0 0.00 383,729 278,123 72.48 382,728 285,790 74.67 03 08 Ingworth & Sparham 27,983 6.66 425,260 239,965 56.43 420,215 258,960 61.63 727,583 9.28 7,913,818 4,789,282 60.52 7,837,491 4,895,456 62.46 01/10/2019 NORWICH DIOCESAN BOARD OF FINANCE LTD DEANERY HISTORY REPORT MONTH September YEAR 2019 SUMMARY PARISH 2017 OUTST. -

Council Tax Rates 2020 - 2021
BRECKLAND COUNCIL NOTICE OF SETTING OF COUNCIL TAX Notice is hereby given that on the twenty seventh day of February 2020 Breckland Council, in accordance with Section 30 of the Local Government Finance Act 1992, approved and duly set for the financial year beginning 1st April 2020 and ending on 31st March 2021 the amounts as set out below as the amount of Council Tax for each category of dwelling in the parts of its area listed below. The amounts below for each parish will be the Council Tax payable for the forthcoming year. COUNCIL TAX RATES 2020 - 2021 A B C D E F G H A B C D E F G H NORFOLK COUNTY 944.34 1101.73 1259.12 1416.51 1731.29 2046.07 2360.85 2833.02 KENNINGHALL 1194.35 1393.40 1592.46 1791.52 2189.63 2587.75 2985.86 3583.04 NORFOLK POLICE & LEXHAM 1182.24 1379.28 1576.32 1773.36 2167.44 2561.52 2955.60 3546.72 175.38 204.61 233.84 263.07 321.53 379.99 438.45 526.14 CRIME COMMISSIONER BRECKLAND 62.52 72.94 83.36 93.78 114.62 135.46 156.30 187.56 LITCHAM 1214.50 1416.91 1619.33 1821.75 2226.58 2631.41 3036.25 3643.49 LONGHAM 1229.13 1433.99 1638.84 1843.70 2253.41 2663.12 3072.83 3687.40 ASHILL 1212.28 1414.33 1616.37 1818.42 2222.51 2626.61 3030.70 3636.84 LOPHAM NORTH 1192.57 1391.33 1590.09 1788.85 2186.37 2583.90 2981.42 3577.70 ATTLEBOROUGH 1284.23 1498.27 1712.31 1926.35 2354.42 2782.50 3210.58 3852.69 LOPHAM SOUTH 1197.11 1396.63 1596.15 1795.67 2194.71 2593.74 2992.78 3591.34 BANHAM 1204.41 1405.14 1605.87 1806.61 2208.08 2609.55 3011.01 3613.22 LYNFORD 1182.24 1379.28 1576.32 1773.36 2167.44 2561.52 2955.60 3546.72 -

Contents of Volume 14 Norwich Marriages 1813-37 (Are Distinguished by Letter Code, Given Below) Those from 1801-13 Have Also Been Transcribed and Have No Code
Norfolk Family History Society Norfolk Marriages 1801-1837 The contents of Volume 14 Norwich Marriages 1813-37 (are distinguished by letter code, given below) those from 1801-13 have also been transcribed and have no code. ASt All Saints Hel St. Helen’s MyM St. Mary in the S&J St. Simon & St. And St. Andrew’s Jam St. James’ Marsh Jude Aug St. Augustine’s Jma St. John McC St. Michael Coslany Ste St. Stephen’s Ben St. Benedict’s Maddermarket McP St. Michael at Plea Swi St. Swithen’s JSe St. John Sepulchre McT St. Michael at Thorn Cle St. Clement’s Erh Earlham St. Mary’s Edm St. Edmund’s JTi St. John Timberhill Pau St. Paul’s Etn Eaton St. Andrew’s Eth St. Etheldreda’s Jul St. Julian’s PHu St. Peter Hungate GCo St. George Colegate Law St. Lawrence’s PMa St. Peter Mancroft Hei Heigham St. GTo St. George Mgt St. Margaret’s PpM St. Peter per Bartholomew Tombland MtO St. Martin at Oak Mountergate Lak Lakenham St. John Gil St. Giles’ MtP St. Martin at Palace PSo St. Peter Southgate the Baptist and All Grg St. Gregory’s MyC St. Mary Coslany Sav St. Saviour’s Saints The 25 Suffolk parishes Ashby Burgh Castle (Nfk 1974) Gisleham Kessingland Mutford Barnby Carlton Colville Gorleston (Nfk 1889) Kirkley Oulton Belton (Nfk 1974) Corton Gunton Knettishall Pakefield Blundeston Cove, North Herringfleet Lound Rushmere Bradwell (Nfk 1974) Fritton (Nfk 1974) Hopton (Nfk 1974) Lowestoft Somerleyton The Norfolk parishes 1 Acle 36 Barton Bendish St Andrew 71 Bodham 106 Burlingham St Edmond 141 Colney 2 Alburgh 37 Barton Bendish St Mary 72 Bodney 107 Burlingham -

Hempton to South Creake Cable Route Hempton, Dunton, Sculthorpe and South Creake Norfolk
Hempton to South Creake Cable Route Hempton, Dunton, Sculthorpe and South Creake Norfolk Programme of Archaeological Recording for Lark Energy CA Project: 660529 CA Report: 15762 SMS and Earthwork Survey Event No: ENF138499 Watching Brief Event No: ENF138543 October 2015 Hempton to South Creake Cable Route Hempton, Dunton, Sculthorpe and South Creake Norfolk Programme of Archaeological Recording CA Project: 660529 CA Report: 15762 Document Control Grid Revision Date Author Checked by Status Reasons for Approved revision by A 15/10/2015 SRJ SCC Internal SCC review This report is confidential to the client. Cotswold Archaeology accepts no responsibility or liability to any third party to whom this report, or any part of it, is made known. Any such party relies upon this report entirely at their own risk. No part of this report may be reproduced by any means without permission. © Cotswold Archaeology © Cotswold Archaeology Hempton to South Creake cable route: Programme of Archaeological Recording CONTENTS SUMMARY ..................................................................................................................... 2 1. INTRODUCTION ................................................................................................ 3 2. ARCHAEOLOGICAL BACKGROUND ................................................................ 4 3. AIMS AND OBJECTIVES ................................................................................... 5 4. METHODOLOGY .............................................................................................. -

Norfolk Vanguard Offshore Wind Farm Consultation Report Appendix 20.3 Socc Stakeholder Mailing List
Norfolk Vanguard Offshore Wind Farm Consultation Report Appendix 20.3 SoCC Stakeholder Mailing List Applicant: Norfolk Vanguard Limited Document Reference: 5.1 Pursuant to APFP Regulation: 5(2)(q) Date: June 2018 Revision: Version 1 Author: BECG Photo: Kentish Flats Offshore Wind Farm This page is intentionally blank. Norfolk Vanguard Offshore Wind Farm Appendices Parish Councils Bacton and Edingthorpe Parish Council Witton and Ridlington Parish Council Brandiston Parish Council Guestwick Parish Council Little Witchingham Parish Council Marsham Parish Council Twyford Parish Council Lexham Parish Council Yaxham Parish Council Whinburgh and Westfield Parish Council Holme Hale Parish Council Bintree Parish Council North Tuddenham Parish Council Colkirk Parish Council Sporle with Palgrave Parish Council Shipdham Parish Council Bradenham Parish Council Paston Parish Council Worstead Parish Council Swanton Abbott Parish Council Alby with Thwaite Parish Council Skeyton Parish Council Melton Constable Parish Council Thurning Parish Council Pudding Norton Parish Council East Ruston Parish Council Hanworth Parish Council Briston Parish Council Kempstone Parish Council Brisley Parish Council Ingworth Parish Council Westwick Parish Council Stibbard Parish Council Themelthorpe Parish Council Burgh and Tuttington Parish Council Blickling Parish Council Oulton Parish Council Wood Dalling Parish Council Salle Parish Council Booton Parish Council Great Witchingham Parish Council Aylsham Town Council Heydon Parish Council Foulsham Parish Council Reepham -

Holme Hale | Norfolk | IP25 7EE STATELY MAGNIFICENCE
Holme Hale | Norfolk | IP25 7EE STATELY MAGNIFICENCE Standing in twelve acres of the most beautiful grounds including a large garden, a small lake and an area of woodland, this breath-taking period country house boasts a grand reception hall and numerous living and entertaining spaces that have to be seen to be believed. Offering 5/6 bedrooms, three en-suite bathrooms and, outside, two garages, a stable and workshop with further detached stores, this substantial property really has it all. Situated a short distance from the village of Holme Hale, the market town of Swaffham is approximately five miles by road, and the Cathedral City of Norwich is approximately twenty miles, with its main line station to London. Opportunities in this part of Norfolk to purchase such an exceptional country house set in what amounts to its own estate are rare, and early viewing is highly recommended. • Picturesque setting for this Beautiful Family Home, hidden from sight in Rural Norfolk Village • Sitting in grounds of just over 12 Acres (12.2 STMS) with Lake, Pond and Wood Areas • Reception Hall, Six Reception Rooms, Kitchen/Breakfast Room, Utility and Laundry Room • Five Bedrooms, Family Bathroom & Three En-Suites • Range of Outbuildings incl. Double Garage, Triple Garage and Workshop and Studio • Total Accommodation including outbuildings extends to 6932 sq.ft • Energy Rating D STATELY MAGNIFICENCE “It once had a railway station on the line between Swaffham and Thetford. The station and goods sheds are now private dwellings and form part of the satellite hamlet of Holme Hale Station Road” Luxury Living Approached via a long graveled drive, the front aspect of Willow House is of flint construction with Gothic arched windows, topped by a remarkable steep-sided tiled roof which curves around at either end to incorporate the fabulous circular features found there. -

Parish Registers and Transcripts in the Norfolk Record Office
Parish Registers and Transcripts in the Norfolk Record Office This list summarises the Norfolk Record Office’s (NRO’s) holdings of parish (Church of England) registers and of transcripts and other copies of them. Parish Registers The NRO holds registers of baptisms, marriages, burials and banns of marriage for most parishes in the Diocese of Norwich (including Suffolk parishes in and near Lowestoft in the deanery of Lothingland) and part of the Diocese of Ely in south-west Norfolk (parishes in the deanery of Fincham and Feltwell). Some Norfolk parish records remain in the churches, especially more recent registers, which may be still in use. In the extreme west of the county, records for parishes in the deanery of Wisbech Lynn Marshland are deposited in the Wisbech and Fenland Museum, whilst Welney parish records are at the Cambridgeshire Record Office. The covering dates of registers in the following list do not conceal any gaps of more than ten years; for the populous urban parishes (such as Great Yarmouth) smaller gaps are indicated. Whenever microfiche or microfilm copies are available they must be used in place of the original registers, some of which are unfit for production. A few parish registers have been digitally photographed and the images are available on computers in the NRO's searchroom. The digital images were produced as a result of partnership projects with other groups and organizations, so we are not able to supply copies of whole registers (either as hard copies or on CD or in any other digital format), although in most cases we have permission to provide printout copies of individual entries. -

Agenda, So As to Save Any Unnecessary Waiting by Members of the Public Attending for Such Applications
Development Committee Please contact: Linda Yarham Please email: [email protected] Direct Dial: 01263 516019 14 February 2018 A meeting of the Development Committee will be held in the Council Chamber at the Council Offices, Holt Road, Cromer on Thursday 22 February 2018 at 9.30am. Coffee will be available for Members at 9.00am and 11.00am when there will be a short break in the meeting. A break of at least 30 minutes will be taken at 1.00pm if the meeting is still in session. Any site inspections will take place on Thursday 15 March 2018. PUBLIC SPEAKING – TELEPHONE REGISTRATION REQUIRED Members of the public who wish to speak on applications are required to register by 9 am on Tuesday 20 February 2018 by telephoning Customer Services on 01263 516150. Please read the information on the procedure for public speaking on our website here or request a copy of “Have Your Say” from Customer Services. Anyone attending this meeting may take photographs, film or audio-record the proceedings and report on the meeting. Anyone wishing to do so must inform the Chairman. If you are a member of the public and you wish to speak, please be aware that you may be filmed or photographed. Emma Denny Democratic Services Manager To: Mrs S Arnold, Mrs A Fitch-Tillett, Mrs A Green, Mrs P Grove-Jones, Mr B Hannah, Mr N Lloyd, Mr N Pearce, Ms M Prior, Mr R Reynolds, Mr P Rice, Mr S Shaw, Mr R Shepherd, Mr B Smith, Mrs V Uprichard Substitutes: Mr D Baker, Mrs S Bütikofer, Mr N Coppack, Mrs A Claussen-Reynolds, Mrs J English, Mr T FitzPatrick, Mr V FitzPatrick, Mr S Hester, Mr M Knowles, Mrs B McGoun, Mrs J Oliver, Miss B Palmer, Mrs G Perry-Warnes, Mr J Punchard, Mr J Rest, Mr E Seward, Mr D Smith, Mr N Smith, Ms K Ward, Mr A Yiasimi All other Members of the Council for information. -
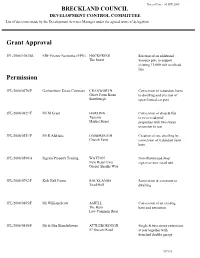
Grant Approval Permission
Date of List - 30 SEP 2008 BRECKLAND COUNCIL DEVELOPMENT CONTROL COMMITTEE List of decisions made by the Development Services Manager under the agreed terms of delegation. Grant Approval 3PL/2008/1092/SU EDF Energy Networks (EPN) HOCKERING Erection of an additional The Street wooden pole to support existing 11,000 volt overhead line Permission 3PL/2008/0276/F Gorhambury Estate Company CRANWORTH Conversion of redundant barns Grove Farm Barns to dwelling and erection of Southburgh open fronted car port 3PL/2008/0421/F Mr M Grant HARLING Conversion of shop & flat Tuscany to two residential Market Street properties with two storey extension to rear 3PL/2008/0551/F Mr R Aldridge OXBOROUGH Creation of one dwelling by Church Farm conversion of redundant farm barn 3PL/2008/0590/A Ingram Property Trading WATTON Non -illuminated shop New Retail Unit sign over new retail unit Gregor Shanks Way 3PL/2008/0792/F Kirk Hall Farms ROCKLANDS Renovation & extension to Toad Hall dwelling 3PL/2008/0805/F Mr William Scott ASHILL Conversion of an existing The Barn barn and extension Low Common Road 3PL/2008/0850/F Mr & Mrs Blanchflower ATTLEBOROUGH Single & two storey extensions 57 Queens Road at rear together with detached double garage DC135 Date of List - 30 SEP 2008 BRECKLAND COUNCIL DEVELOPMENT CONTROL COMMITTEE List of decisions made by the Development Services Manager under the agreed terms of delegation. Permission 3PL/2008/0889/F Mr G Tweed SAHAM TONEY Change of use of 94 Richmond Road existing building to farm shop 3PL/2008/0915/F Mr M Gore DEREHAM -

Doc - Complete
Date of List - 11 AUG 2016 BRECKLAND COUNCIL PLANNING COMMITTEE List of decisions made by the Breckland Council under the agreed terms of delegation. DOC - COMPLETE 3DC/2011/0142/DOC Mr J F Rice GREAT DUNHAM Discharge of conditions 5, 6, Riceland 11, 12, 13 & 14 on pp Litcham Road 3PL/2011/0539/F 3DC/2013/0117/DOC Mr David Sayer SPARHAM Discharge of conditions 4, 8, Well Lane 9, 11, 13 & 20 of pp 3PL/2012/0050/F 3DC/2016/0101/DOC Wrights Produce Ltd BRIDGHAM Conversion of existing open Barn Farm cattle enclosures to seven The Street pig finishing barns for the rearing of production pigs 3DC/2011/0143/DOC Mr M & Mrs S Daisley FRANSHAM Discharge of conditions 3 & 4 Plot adjacent The Manor of PP 3PL/2010/0830/D Main Road 3DC/2015/0059/DOC Mr Matthew Newbury KENNINGHALL Discharge of conditions 4,8 & Land East of Memorial Hall 10 on 3PL/2011/1226/F School Close 3DC/2016/0057/DOC Miss Elizabeth Hodder THETFORD Confirmation that conditions 2 Nickerson Yard have been discharged on Castle Street 3PL/2007/1554/F & 3PL/2007/1613/LB 3DC/2012/0060/DOC J D Alston (Estates) KENNINGHALL Discharge of conditions 3, 4, Guiltcross Dairy 5 and 6 of pp 3PL/2011/1307/F Garboldisham Road 3DC/2016/0071/DOC Mr Robert Williams OLD BUCKENHAM Discharge of conditions 3 & The Barn, Orchard House 12 on 3PL/2015/1410/F Hargham Road DC135 (2016) Date of List - 11 AUG 2016 BRECKLAND COUNCIL PLANNING COMMITTEE List of decisions made by the Breckland Council under the agreed terms of delegation. -

Winter/Spring 2005 ● Issue No.9
Winter/Spring 2005 ● Issue No.9 HOLKHAM NEWSLETTER HERE might be a perception that all the changes we have seen at Holkham have taken place only in the last five or six years.This process might have been more rapid in that time, but that only Treflects the speed of change in the world in general. Change has always been with us. Mick Thompson, recently retired from the Hall, recalls how he helped to modernise four milking parlours on the Estate.Well, how many have we now? None: this is change brought on by economic circumstances. I started farming at Burnham Norton, 36 years ago, with 11 men. By the time the Holkham Farming Company took over, that was down to only three, again a reflection of changed economic circumstances, and a necessary adaptation to new farming practices. Now, seven men look after 5,000 acres. A good example of adaptation is the transformation of the 18th century Triumphal Arch into a chic annexe of The Victoria Hotel, thus turning a long-standing liability to economic advantage, and moreover, making use of a perfectly sound building. I never see the point of new build if existing buildings can be satisfactorily adapted without compromising their character. But with change, it is important to retain and value the best of the past. Nicholas Hills writes about the restoration of the family monuments at Tittleshall Church, to which the Estate contributed a large part of the £70,000 costs.We could have saved our money, but then part of the history of this great Estate would have been lost. -

NOTICE of ELECTION Election of Parish Councillors
NOTICE OF ELECTION Breckland Election of Parish Councillors for the Parishes listed below Number of Number of Parish Parish Parishes Parishes Councillors to Councillors to be elected be elected Parish of Ashill Nine (9) Parish of Little Dunham Seven (7) Parish of Banham Nine (9) Parish of Little Ellingham Five (5) Parish of Bawdeswell Seven (7) Parish of Longham Seven (7) Parish of Beachamwell Seven (7) Parish of Lyng Seven (7) Parish of Beeston with Bittering Seven (7) Parish of Mattishall Nine (9) Parish of Beetley Seven (7) Parish of Merton Five (5) Parish of Besthorpe Seven (7) Parish of Mileham Seven (7) Parish of Billingford Seven (7) Parish of Mundford Nine (9) Parish of Bintree Seven (7) Parish of Narborough Seven (7) Parish of Blo` Norton Five (5) Parish of New Buckenham Seven (7) Parish of Bradenham Seven (7) Parish of Necton Nine (9) Parish of Brettenham and Seven (7) Parish of North Elmham Eleven(11) Kilverstone Parish of Bridgham Five (5) Parish of North Lopham Seven (7) Parish of Brisley Seven (7) Parish of North Pickenham Seven (7) Parish of Carbrooke Nine (9) Parish of North Tuddenham Seven (7) Parish of Caston Seven (7) Parish of Old Buckenham Eleven(11) Parish of Cockley Cley Five (5) Parish of Ovington Five (5) Parish of Colkirk Seven (7) Parish of Oxborough Five (5) Parish of Cranworth Seven (7) Parish of Quidenham Seven (7) Parish of Croxton Five (5) Parish of Rocklands Seven (7) Parish of East Tuddenham Seven (7) Parish of Rougham Seven (7) Parish of Elsing Seven (7) Parish of Roudham and Larling Seven (7) Parish