Blyttia 34-4.Pdf
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Fylkesmannens Kommunebilde Evje Og Hornnes Kommune 2013
Fylkesmannens kommunebilde Evje og Hornnes kommune 2013 Innhold 1 Innledning 3 2 Planlegging og styring av kommunen 4 2.1 Demografi 4 2.2 Likestilling 5 2.3 Økonomi og økonomistyring 8 2.4 Bosetting og kvalifisering av flyktninger 12 3 Tjenesteyting og velferdsproduksjon 14 3.1 Barnehage 14 3.2 Tidlig innsats 18 3.3 Utdanning 19 3.4 Vergemål 25 3.5 Byggesak 26 3.6 Folkehelse og levekår 27 3.7 Barneverntjenesten 29 3.8 Helsesøster- og jordmortjeneste 32 3.9 Helsetjenester 33 3.10 Tjenester i sykehjem 35 3.11 Velferdsprofil 37 3.12 Arealplanlegging 39 3.13 Forurensning 40 3.14 Landbruk 41 Side: 2 - Fylkesmannen i Aust-Agder, kommunebilde 2013 Evje og Hornnes 1 Innledning Kommunebilde, som Fylkesmannen utarbeider for alle 15 kommuner i Aust-Agder, er et viktig element i styringsdialogen mellom kommunene og Fylkesmannen. Det gir ikke et fullstendig utfyllende bilde av kommunens virksomhet, men gjelder de felt der Fylkesmannen har oppdrag og dialog mot kommunene. Det er et dokument som peker på utfordringer og handlingsrom. Fylkesmannen er opptatt av at kommunene kan løse sine oppgaver best mulig og ha politisk og administrativt handlingsrom. Det er viktig at kommunene har et best mulig beslutningsgrunnlag for de valg de tar. Blant annet derfor er kommunebilde et viktig dokument. Kommunebilde er inndelt i tre 1. Innledning 2. Planlegging og styring av kommunen 3. Tjenesteyting og velferdsproduksjon Under hvert punkt har vi oppsummert Fylkesmannens vurdering av utfordringspunkter for kommunen. Dette er punkter vi anbefaler kommunen å legge til grunn i videre kommunalplanlegging. Kommunebilde er et godt utgangspunkt for å få et makrobilde av kommunen og bør brukes aktivt i kommunen – ikke minst av de folkevalgte. -

Alkohol 2010
Vest-Agder-museet a Årbok 2010 l ko h ol alkohol ”Drikk feller ungdommen i hans blomst, mannen i hans kraft og oldingen i hans svakhet. Den knuser farens hjerte, gjennomborer morens ømme barm, slukker den naturlige hengivenhet, kveler ektekjærligheten, utsletter den sønnlige frykt, tilintetgjør foreldrenes håp og legger deres grå hår med sorg i graven. Dens virkning er svakhet istedenfor kraft, sykdom istedenfor sunnhet, død istedenfor liv.” 2010 Gleder og sorger 9 7 8 8 2 9 1 1 7 8 2 0 2 2010 ÅRBOK VEST-AGDER-MUSEET 2010 Utgitt av Vest-Agder-museet IKS © Redaktør Thor Gunnar Hansen Redaksjon Tale Christiansen og Thorunn Lunde Design Per Grimsgaard Layout og trykk Kai Hansen Trykkeri, Kristiansand Opplag 1000 ISBN: 978-82-91178-20-2 Vest-Agder-Museet IKS Odderøya 17 4610 Kristiansand www.vestagdermuseet.no Sitatet på omslaget er hentet fra den amerikanske juristen og forfatteren Robert Green Ingersoll (1833 - 1899). Fotograf Ragnvald Pettersen har tatt bildene som er på omslag og forsats i årboka. Han overtok familiebedriften i 1884 og drev den til 1942. Atelieret hadde fra 1884 adresse ”vis a vis Lundhs have” i Farsund. Fotografiene viser yngre menn på besøk i fotoatelieret og gjengitt i ledige, kameratslige situasjoner. Motivene skiller seg fra typiske visittkort som var populære fra andre halvdel av 1800-tallet. De var heller ikke skrytebilder egnet til familiealbumet. Det var vanlig med bruk av ulike rekvisitter i fotoatelierene. Kanskje kan motivene være ment som ungdommelige provokasjoner i en tid med sterk totalavholdsbevegelse? Carl Pettersen, Ragnvalds far, introduserte visittkortet i Norge i 1861. Fotografiene eies av Lyngdal Kultursenter KF. -

140914 Arrangørdagbok Gimletroll 2014-15.Xlsx
Arrangørdagbok for Gimletroll IK 2014‐2015 Tid Ansvarlige Kampnr Respin Avdeling Bane Hjemmelag Bortelag Ansvarlige Obs TIRSDAG 23.09.2014 1900 55K0503007 1195139 5. divisjon Kvinner Avd. 2 Agder Aquarama Gimletroll AK 28 3 Damelaget Damelaget ONSDAG 24.09.2014 2000 55J1304001 1168324 Jenter 13 Avdeling B 2 Havlimyrhallen Gimletroll 2 Fløy 2 Jenter 13 Jenter 13 ONSDAG 08.10.2014 1700 Leif E 55HU‐01001 1134590 Håndball for utviklingshemmede Agder Odderneshallen Grane Arendal 2 Vindbjart v 1740 Leif E 55HU‐01002 1134591 Håndball for utviklingshemmede Agder Odderneshallen MHI Grane Arendal 1 Leif E 1820 Leif E 55HU‐01004 1134593 Håndball for utviklingshemmede Agder Odderneshallen Gimletroll Grane Arendal 2 Leif E 1900 Leif E 55HU‐01003 1134592 Håndball for utviklingshemmede Agder Odderneshallen Vindbjart Våg Leif E 1940 Leif E 55HU‐01005 1134594 Håndball for utviklingshemmede Agder Odderneshallen Greipstad MHI Leif E 2020 Leif E 55HU‐01006 1134595 Håndball for utviklingshemmede Agder Odderneshallen Grane Arendal 1 Imås Leif E 2100 Leif E 55HU‐01008 1134597 Håndball for utviklingshemmede Agder Odderneshallen Imås Gimletroll Leif E 2140 Leif E 55HU‐01007 1134596 Håndball for utviklingshemmede Agder Odderneshallen Våg Greipstad Leif E SØNDAG 12.10.2014 1200 Jenter 12 55J1205008 1165085 Jenter 12 Avdeling 5 Havlimyrhallen Gimletroll 3 Torridal * Jenter 12 1300 Jenter 13 55J1304020 1168343 Jenter 13 Avdeling B 2 Havlimyrhallen Gimletroll 2 Kristiansand 2 Jenter 13 1400 55J1401012 1195338 Jenter 14 Avdeling A Agder Havlimyrhallen Gimletroll Strømmen‐Glimt/Hisøy -

Sluttrapport Fra Forvaltningskontroll Landbruk Evje Og Hornnes Og Iveland
Landbruksavdelingen Sluttrapport fra kontroll av forvaltningen av utvalgte tilskuddsordninger i landbruket – 2016 Kontrollert Evje og Hornnes og Iveland Kontrollpersoner fra Hans Bach- enhet kommuner Fylkesmannen i Evensen og Rolf Aust- og Vest-Agder Inge Pettersen Kontrollomfang Dato for sluttrapport 01.12.2016 -Avløsning ved sykdom og fødsel mv. - Tilskudd til spesielle miljøtiltak i landbruket (SMIL). Kommune Kontaktperson Evje og Hornnes og Iveland kommune Inge Eftevand Rapportens innhold Rapporten er utarbeidet etter gjennomført forvaltningskontroll i Iveland og Evje og Hornnes kommune innen ordningene med avløsning ved sykdom og fødsel mv. og tilskudd til spesielle miljøtiltak i landbruket (SMIL). Rapporten er utarbeidet som 1 rapport til begge kommunene siden det er felles forvaltning på de aktuelle landbrukstilskuddene. Den beskriver eventuelle avvik og merknader som ble avdekket innen de kontrollerte områdene. Avvik er mangel på oppfyllelse av krav fastsatt i lov, forskrift og/eller rundskriv. Merknad er forhold som ikke er i strid med krav, men der Fylkesmannen finner grunn til å påpeke behov for forbedring. Bakgrunn for kontrollen Fylkesmannen har det regionale ansvar for å sikre at statlige midler brukes og forvaltes i samsvar med forutsetningene. Forvaltningskontrollen har hjemmelsgrunnlag i Økonomireglementet i staten (§ 15) og bakgrunn i fullmaktsbrev fra Landbruksdirektoratet for 2016. Ved å avdekke eventuelle avvik og brudd på regelverk og rutiner vil dette kunne hjelpe kommunen fram mot en sikrere, bedre og mer effektiv forvaltning av tilskuddsordningene. Gjennomføring og omfang Kontrollen ble gjennomført som en dokumentkontroll. Fylkesmannen gjennomgikk forhåndsvalgte saker som kommunene hadde oversendt. Dette gjaldt 3 saker om tilskudd til avløsning til sykdom og fødsel mv. og 4 saker tilknyttet SMIL ordningen. -

Folketeljing 1900 for 0936 Hornnes Digitalarkivet
Folketeljing 1900 for 0936 Hornnes Digitalarkivet 25.09.2014 Utskrift frå Digitalarkivet, Arkivverket si teneste for publisering av kjelder på internett: http://digitalarkivet.no Digitalarkivet - Arkivverket Innhald Løpande liste .................................. 9 Førenamnsregister ........................ 51 Etternamnsregister ........................ 63 Fødestadregister ............................ 75 Bustadregister ............................... 87 4 Folketeljingar i Noreg Det er halde folketeljingar i Noreg i 1769, 1801, 1815, 1825, 1835, 1845, 1855, 1865, 1870 (i nokre byar), 1875, 1885 (i byane), 1891, 1900, 1910, 1920, 1930, 1946, 1950, 1960, 1970, 1980, 1990 og 2001. Av teljingane før 1865 er berre ho frå 1801 nominativ, dvs. ho listar enkeltpersonar ved namn. Teljingane i 1769 og 1815-55 er numeriske, men med namnelistar i grunnlagsmateriale for nokre prestegjeld. Statistikklova i 1907 la sterke restriksjonar på bruken av nyare teljingar. Etter lov om offisiell statistikk og Statistisk Sentralbyrå (statistikklova) frå 1989 skal desse teljingane ikkje frigjevast før etter 100 år. 1910-teljinga blei difor frigjeven 1. desember 2010. Folketeljingane er avleverte til Arkivverket. Riksarkivet har originalane frå teljingane i 1769, 1801, 1815-1865, 1870, 1891, 1910, 1930, 1950, 1970 og 1980, mens statsarkiva har originalane til teljingane i 1875, 1885, 1900, 1920, 1946 og 1960 for sine distrikt. Folketeljinga 3. desember 1900 Ved kgl. res. 8. august 1900 blei det bestemt å halde ei "almindelig Folketælling" som skulle gje ei detaljert oversikt over befolkninga i Noreg natta mellom 2. og 3. desember 1900. På kvar bustad skulle alle personar til stades førast i teljingslista, med særskild markering ("mt") av dei som var mellombels til stades (på besøk osb.) på teljingstidspunktet. I tillegg skulle alle faste bebuarar som var fråverande (på reise, til sjøs osb.) frå bustaden på teljingstidspunktet, også førast i lista, men merkast som fråverande ("f"). -

Evje Og Hornnes Kommune (PDF)
E3AF, ; Evje og Hornnes kommune ' (WLIDERHSDEP/- Rådmann/Administrasjon Arkivkock: 7 z p(( Administrativt vedtak aor( ocil?. — Sak nr. 26/11 Barne-, likestillings- og inkluderingsdepartementet Postboks 8036 Dep 0030 OSLO Vår ref:Saksbeh:Arkivkode: Dato: 2010/445-11Line Håberg Løvdal 2 37 93 23 22F30 28.04.2011 E-post: [email protected] Høringsuttale: forslag til bosettingsordning for flyktninger med mål om raskere bosetting Høringsuttale fra Evje og Hornnes kommune er behandlet av rådmannen i administrativt vedtak. I den forbindelse er det gjennomført en intern høring der flyktningtjenesten, leder for NAV Evje og Hornnes, rektor ved voksenopplæringen og leder for Helse og familie har kommet med innspill til kommunens høringsuttale. Ka ittel VI forsla til endrin er i da ens ordnin . Målsettinger for en mer målrettet ordning. Evje og Hornnes kommune støtter målet om at flyktninger skal være bosatt i en kommune senest 6 måneder etter at det er fattet vedtak om opphold, og at enslige mindreårige skal være bosatt innen 3 måneder etter vedtak. Pkt 4. Dersom kommunene i et ftlke ikke blir enige, avgjør ftlket fordelingen. Fylkesmannen gis da rett til å tilvise flyktninger til kommunene som plikter å ta imot den enkelte flyktning. Evje og Hornnes kommune mener at bosetting av flyktninger i størst mulig grad bør baseres på kommunale beslutninger og kommunal frivillighet. Kommunal selvråderett bør være førende. Kommunen er imot høringsforslaget om at Fylkesmannen gis tilvisningsrett overfor kommunene når det gjelder bosetting. Kommunene bør i stedet oppfordres til å ta ansvar for bosetting av flyktninger. Pkt. 5. Fylkesmannens tilvisingsrett skal bare brukes dersom staten anser det som nødvendig for å klare å bosette det antall flyktninger det er behov for. -

MENIGHETEN SAMFUNDET, DET ALMINDELIGE SAMFUND OG ANDRE STERKTTROENDE © 2021 Jonas Yassin Iversen
De gamle stier Jonas Yassin Iversen De gamle stier HISTORIEN OM MENIGHETEN SAMFUNDET, DET ALMINDELIGE SAMFUND OG ANDRE STERKTTROENDE © 2021 Jonas Yassin Iversen Dette verket omfattes av bestemmelsene i Lov om opphavsretten til åndsverk m.v. av 1961. Verket utgis Open Access under betingelsene i Creative Commons-lisensen CC BY-NC-ND 4.0. Denne lisensen lar andre kopiere, distribuere og spre verket i hvilket som helst medium eller format, under forutsetning av at det oppgis korrekt kreditering og lenke til lisens. Dette kan gjøres på enhver rimelig måte, men uten at det kan forstås slik at lisensgiver bifaller deg eller din bruk av verket. Materialet kan ikke benyttes til kommersielle formål. Dersom du remixer, bearbeider eller bygger på materialet, kan du ikke distribuere det endrede materialet. Lisensvilkår: https://creativecommons.org/licenses/by-nc-nd/4.0/deed.no Boka er utgitt med støtte fra Fakultet for lærerutdanning og pedagogikk, Høgskolen i Innlandet, Høgskolen i Innlandets publiseringsfond, Fritt Ord, Dalane folkemuseum og forskergruppen DivE ved Høgskolen i Innlandet. Denne boka er utgivelse nr. 41 i skriftserien Kyrkjefag Profil. Dette er en fagfellevurdert monografi. ISSN print: 1502–7929 ISSN online: 2703-8076 ISBN trykt bok: 978-82-02-71975-3 ISBN PDF: 978-82-02-71974-6 ISBN EPUB: 978-82-02-72051-3 ISBN HTML: 978-82-02-72052-0 ISBN XML: 978-82-02-72054-4 DOI: https://doi.org/10.23865/noasp.131 Omslagsdesign: Cappelen Damm AS Omslagsbilde: Bilde av Det Almindelige Samfunds kirke på Krossmoen utenfor Egersund, med bl.a. forstander Theodor Ege og Kristine Skipstad i forgrunnen. -

Søknader Fra Eksterne / Diverse Vedlegg
Søknader fra eksterne / diverse vedlegg Budsjett for kontrollarbeidet i Evje og Hornnes kommune 2021.......................... Side 2 - 5 Budsjett for 2021 – Agder Sekretariatet................................................................ Side 6 - 9 Otra IL – Søknad om investering og finansiering av løypemaskin.......................... Side 10 Soknerådets søknad om kommunal bevilgning for 2021....................................... Side 11 - 14 Tilskudd til Senter mot seksuelle overgrep Agder SMSO....................................... Side 15 - 18 Søknad om driftstilskudd 2021 – ARKIVET freds- og menneskerettighetssenter (Stiftelsen Arkivet)...................... Side 19 – 21 SEIF - søknad om driftsmidler 2021........................................................................ Side 22 – 24 Stiftelsen Amathea – søknad om driftstilskudd...................................................... Side 25 - 26 UTSKRIFT AV MØTEBOK EVJE OG HORNNES KOMMUNE – KONTROLLUTVALGET Onsdag 28. oktober 2020 SAK 11/20 BUDSJETT FOR KONTROLLARBEIDET I EVJE OG HORNNES KOMMUNE 2021 Fast godtgjørelse på kr. 36.857 til leder var ikke medtatt i opprinnelig budsjettforslag. Utvalget la inn denne posten i budsjettet under behandlingen. Kontrollutvalget fattet følgende vedtak: 1. Kontrollutvalget tilrår en budsjettramme for kontrollarbeidet i Evje og Hornnes kommune for 2021 på kr. 1.212.957. 2. Budsjettforslaget skal følge formannskapets behandling og innstilling til kommunestyret vedr. budsjett 2021 Tabell: Budsjett for kontrollarbeidet i Evje og Hornnes kommune -

Greipstad Pistolklubb Rismyrvegen 61 4645 Nodeland Telefon: 38182410
Greipstad Pistolklubb Stevne: 1M Milfelt Startkontingent: 100,00 Rismyrvegen 61 Dato: 25.05.2014 Maks 108 4645 Nodeland Stevnenummer: 1411018 poengsum: Telefon: 38182410 Bane: Sandrip Premieandel: 28,00 Epost: Stevneleder: Jan Ivar Ingebrethsen Til bestemann: 3,00 [email protected] Jan Ivar Ingebrethsen, Jarl-Edvind KM Dommer(e): Hjemmeside: Edvardsen, Trond Arild Nodeland Mesterskap: Vest- www.greipstadpistolklubb.no Agder Deltakere: 51 Klasse A 1 2 3 4 5 6 7 8 9 10 InnerX Omsk Sum Oppr. Mrk. Premie 1 Dag Liestøl Vennesla PK 9 11 12 12 11 9 11 11 11 8 32 105 A G 276* 2 Roy Baumann Kristiansand PK 9 9 12 12 11 10 11 11 9 10 34 104 A G 98 3 Pål Supersaxo Kristiansand PK 9 11 10 12 8 9 11 11 11 8 27 22 100 B S 52 4 Trond Arild Nodeland M Greipstad Pistolklubb 9 11 12 10 9 7 11 9 11 10 27 99 B S 35 5 Ingunn Edvardsen K Greipstad Pistolklubb 9 11 12 10 9 10 7 11 9 10 36 98 B S 0 6 Per Rune Magnussen Sola PK 9 8 8 12 11 8 7 11 11 10 28 95 B S 0 7 Kjell Eivind Lund Kvinesdal PK 9 11 8 10 6 8 10 11 9 10 21 92 B S 0 8 Runar Magnussen Sola PK 9 11 12 10 6 10 8 9 11 5 27 91 B S 0 9 Knut Stamland Porsgrunn Pistolklubb 9 7 10 10 7 6 9 11 9 4 19 82 C B 0 10 Morten Stamland Norsk Politiidrettsforbund 9 6 4 4 9 0 9 2 3 5 15 51 D 0 11 Jan B. -

«Et Boligområde Blir Aldri Helt Ferdig» Fortetting Med Strøksanalyser Som Verktøy
«Et boligområde blir aldri helt ferdig» Fortetting med strøksanalyser som verktøy Justvik Fagerholt Hånes Sødal Gimlekollen Kongsgård Vige Eg Strømme Lund Søm Grim Kuholmen Korsvik Odderhei Dvergnes Flekkerøya Andøya Forord Rapporten bygger på et arbeid som har vært utført ved Plan- og bygningsetaten i Kristiansand kommune. Prosjektet har gått over mange år, hele tiden på siden av og underordnet de ordinære oppgavene. Samtidig har det gitt innsikt og kunnskap som har kommet til nytte i det løpende arbeidet med planer og byggesaker. Vest-Agder fylkeskommune har støttet prosjektet. Kristiansand 21. januar 2013 Gunnar Stavrum, Plan- og bygningssjef FORTETTING I BOLIGOMRÅDER MED STRØKSANALYSER SOM VERKTØY RAPPORT Innholdsfortegnelse Innledning ................................................................................................................................................ 3 Strøksanalyser ......................................................................................................................................... 7 Bevaring og Vern ................................................................................................................................... 16 Fortetting................................................................................................................................................ 23 Konklusjon ............................................................................................................................................. 34 Litteraturliste ......................................................................................................................................... -
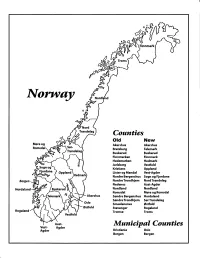
Norway Maps.Pdf
Finnmark lVorwny Trondelag Counties old New Akershus Akershus Bratsberg Telemark Buskerud Buskerud Finnmarken Finnmark Hedemarken Hedmark Jarlsberg Vestfold Kristians Oppland Oppland Lister og Mandal Vest-Agder Nordre Bergenshus Sogn og Fjordane NordreTrondhjem NordTrondelag Nedenes Aust-Agder Nordland Nordland Romsdal Mgre og Romsdal Akershus Sgndre Bergenshus Hordaland SsndreTrondhjem SorTrondelag Oslo Smaalenenes Ostfold Ostfold Stavanger Rogaland Rogaland Tromso Troms Vestfold Aust- Municipal Counties Vest- Agder Agder Kristiania Oslo Bergen Bergen A Feiring ((r Hurdal /\Langset /, \ Alc,ersltus Eidsvoll og Oslo Bjorke \ \\ r- -// Nannestad Heni ,Gi'erdrum Lilliestrom {", {udenes\ ,/\ Aurpkog )Y' ,\ I :' 'lv- '/t:ri \r*r/ t *) I ,I odfltisard l,t Enebakk Nordbv { Frog ) L-[--h il 6- As xrarctaa bak I { ':-\ I Vestby Hvitsten 'ca{a", 'l 4 ,- Holen :\saner Aust-Agder Valle 6rrl-1\ r--- Hylestad l- Austad 7/ Sandes - ,t'r ,'-' aa Gjovdal -.\. '\.-- ! Tovdal ,V-u-/ Vegarshei I *r""i'9^ _t Amli Risor -Ytre ,/ Ssndel Holt vtdestran \ -'ar^/Froland lveland ffi Bergen E- o;l'.t r 'aa*rrra- I t T ]***,,.\ I BYFJORDEN srl ffitt\ --- I 9r Mulen €'r A I t \ t Krohnengen Nordnest Fjellet \ XfC KORSKIRKEN t Nostet "r. I igvono i Leitet I Dokken DOMKIRKEN Dar;sird\ W \ - cyu8npris Lappen LAKSEVAG 'I Uran ,t' \ r-r -,4egry,*T-* \ ilJ]' *.,, Legdene ,rrf\t llruoAs \ o Kirstianborg ,'t? FYLLINGSDALEN {lil};h;h';ltft t)\l/ I t ,a o ff ui Mannasverkl , I t I t /_l-, Fjosanger I ,r-tJ 1r,7" N.fl.nd I r\a ,, , i, I, ,- Buslr,rrud I I N-(f i t\torbo \) l,/ Nes l-t' I J Viker -- l^ -- ---{a - tc')rt"- i Vtre Adal -o-r Uvdal ) Hgnefoss Y':TTS Tryistr-and Sigdal Veggli oJ Rollag ,y Lvnqdal J .--l/Tranbv *\, Frogn6r.tr Flesberg ; \. -

Gustav Vigeland
IBSEN COURIER This newsletter and others can be viewed/printed in color at: http://www.llund.com/newsletters/ Visit our web page at: http://www.llund.com/sons.htm SONS OF NORWAY September 2011 HENRIK IBSEN LODGE No. 3-393 Officers 2011 elcome back accepted my invitation and will be our W everyone!! guest at our November meeting. They will President:* Kenneth Johnson present a forty minute presentation on [email protected] Well it was a hot Norwegian immigration. I’m sure all our Vice President:* summer and I hope families can relate. Helen Timony everyone made it Secretary:* Anita Kindberg through okay. We As we move through the next year I want Treasurer:* Presidents Message are now beginning to remind all our officers of their individ- Kristine Holvik our 2011-2012 busi- ual responsibilities and that we need to Financial Secretary:* ness season but before I mention the continue with the idea of having a chair- Cynthia Biancospino things I think you should know I first person for all our events. Counselor:* Asbjørn Lageraaen want to say thanks to those who sup- Social Director: ported our lodge and of course our sum- As previously mentioned we will not have Darlene Kern mer get-to-gether events. a regular September meeting at the Assistants: Crystal NiFong & Oriya Bartow church but rather when we will be at Editor: A special thanks to Asbjørn our past LOV. Our next regular meeting will be Lawrence Lund President and Counselor for covering for October 21st and I hope that we will all Sports & Recreation Director: me and our VP Helen when we were un- eat together again.