Palaeoecological Studies of Selected Mires in the Craven District of West Yorkshire with Special Reference to the Late Devensian Period and the Ulmus Decline
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Shrub List for Brighton 2010
Shrub List For Brighton 2010 Large Shrubs 10’ -20’ Tall by 6’ – 25’ wide Acer ginnala Amur Maple Acer tataricum Tatarian Maple (better than Amur Maple) Acer grandidentatum Bigtooth Maple Amelanchier alnifolia Saskatoon Serviceberry Amelanchier canadensis Shadblow Serviceberry Caragana arborescens Siberian Peashrub Cercocarpus ledifolius Mountain Mahogany Cotoneaster lucidus Peking Cotoneaster Cowania mexicana Quince Bush, Cliffrose Crataefus ambigua Russian Hawthorn Forestiera neomexicana New Mexican Privet Hippophae rhamnoides Sea Buckthorn Juniperus species Juniper Kolkwitzia amabilis Beauty Bush Pinus mugo Mugo Pine species Prunus americana American Plum Prunus virginiana ‘Shubert’ Canada Red Chokecherry Ptelea trifoliata Wafer Ash or Hop tree Quercus gambelii Gambel Oak Rhus typhina Staghorn Sumac Robinia neomexicana New Mexico Locust Sambucus species Elders Shepherdia argentea Buffaloberry Syringa vulgaris Common Lilac Viburnum lantana Wayfaring Tree, Viburnum Medium Size Shrubs >10’ high by >8’ wide Amorpha fruticosa False Indigo Atriplex canescens Fourwing Saltbush Buddleia davidii Butterfly Bush Cercocarpus montanus Mountain Mahogany Chamaebatiaria millefolium Fernbush Chrysothamnus nauseosus Rubber Rabbitbrush Cornus sericea Redtwig Dogwood Cotinus coggygria Smoke Tree Cotoneaster species Cotoneaster Cytisus scoparius ‘Moonlight’ Moonlight Broom Euonymus alatus Burning Bush Forsythia x intermedia Forsythia Hibiscus syriacus Rose-of-Sharon Juniperus species Juniper Ligustrum vulgare Privet Lonicera species Honeysuckle Mahonia aquifolium Oregon Grape Holly Philadelphus species Mockorange Pyracantha coccinea Firethorn Physocarpus opulifolius Common Ninebark Prunus besseyi Western Sand Cherry Pyracantha coccinea species Firethorn Rhamnus frangula Glossy Buckthorn Ribes species Currant Sambucus species Elder Spiraea x vanhouttei Vanhouttei Spirea Symphoricarpos albus Snowberry Syringa meyeri „Palibin‟ Dwarf Korean Lilac Syringa patula „Miss Kim‟ Dwarf Lilac Viburnum species (dozens of different types) Small Size Shrubs > 5’ tall by >6. -

Find out More About the Three Peaks Project At
The Yorkshire Three Peaks walk Distance: 39km (24 miles) Parking: Horton car park ( BD24 0HF, SD 807 724) Other transport: Horton train station on the Settle to Carlisle line is close to the start Toilets: Horton car park Refreshments: pubs and café in Horton, Station Inn at Ribblehead and the Old Hill Inn in Chapel-le-dale This is a major challenge walk which is long and involves over 1600m (5000 feet) of climbing over the Three Peaks of Pen-y-ghent, Whernside and Ingleborough. There is one section on road, but the paths are good. You do need to be able to navigate and cope with conditions in the high fells. Route description 1. Walk south out of the village passing the Golden Lion pub and church and cross a small stream. Then turn left up a minor tarmac road. Follow this up towards Brackenbottom and just before reaching some buildings take a footpath on your left signed to Pen-y-ghent. 2. Climb steadily up through fields with Pen-y-ghent ahead of you. The final section of the route to the summit is steeper for a while before reaching the trig point and shelter. 3. Cross the wall at the summit and follow the clear path heading roughly north. This zig zags down, passing the gash of Hunt Pot, to reach the head of a walled lane. 4. Carry straight on to follow the new path over Whitber Hill to reach a clear track. Turn right and follow this for 1.5km (1 mile) and then take the path on the left towards Birkwith cave. -

Dent to Aye Gill Pike This Rewarding Walk Has Spectacular Views of the Howgill Fells (Below), the Three Peaks of Yorkshire, and Dentdale
Dent to Aye Gill Pike This rewarding walk has spectacular views of the Howgill Fells (below), the Three Peaks of Yorkshire, and Dentdale. Start: Dent car park (GPS: SD 703 871) Distance: 10.5km (6.5 miles) Highest point: 556m (Aye Gill Pike summit) Time: Allow 3 to 4 hours Grade: Difficult Notes: This route has sections with no obvious path, so a map and navigational skills are required. There are short sections on quiet roads where care should be taken, and the route can be boggy in places. There are toilets, a village shop, café and pubs in Dent. Other options: For a more challenging experience, follow this route in the opposite direction, beginning with the very steep climb up to Aye Gill Pike. 1. (GPS: SD 704 870) Turn left out of the car park and walk through the cobbled village centre. Leave the village on this road and pick up the riverside path for the Dales Way on your left, just before the river Dee. Follow this path until you reach the tarmac road. 2. (GPS: SD 700 873) Turn right along the road and pick up the Dales Way again on your right, continuing along the river to the road at Barth Bridge. 3. (GPS: SD 694 878) At the road turn right. Walk along here for a short distance (approx 150m) and turn up the small tarmac lane on your right. Continue along this quiet lane for approximately 1km, until reaching Lunds Farm. 4. (GPS: SD 694 887) At the entrance to Lunds Farm turn left and continue through the cobbled yard. -

Exploitation of Sea Buckthorn Fruit for Novel Fermented Foods Production: a Review
processes Review Exploitation of Sea Buckthorn Fruit for Novel Fermented Foods Production: A Review Svetlana Schubertová * , Zuzana Krepsová,Lívia Janotková , Marianna Potoˇcˇnáková and František Kreps * Department of Food Technology, Faculty of Chemical and Food Technology, Slovak University of Technology in Bratislava, Radlinského 9, 812 37 Bratislava, Slovakia; [email protected] (Z.K.); [email protected] (L.J.); [email protected] (M.P.) * Correspondence: [email protected] (S.S.); [email protected] (F.K.) Abstract: Sea buckthorn fruit is abundant with essential nutrients and bioactive substances, yet it remains less sought after. Therefore, it is valuable to explore new ways of sea buckthorn fruit processing, which can boost consumer acceptance of sea buckthorn fruit and also lead to formulation of new functional foods. In the presented review, we summarize studies focused on development of foods utilizing sea buckthorn fruit or its components and bacterial food cultures. Firstly, we discuss the impact of malolactic fermentation on content and profile of organic acids and polyphenols of sea buckthorn fruit juice. During this process, changes in antioxidant and sensory properties are considerable. Secondly, we address the role of sea buckthorn fruit and its components in formulating novel probiotic dairy and non-dairy products. In this regard, a synergic effect of prebiotic material and probiotic bacteria against pathogens is distinguished. Overall, the potential of sea buckthorn fruit as a botanical ingredient for application in novel foods is highlighted. Keywords: lactic acid bacteria; malolactic fermentation; novel food; probiotic; sea buckthorn fruit Citation: Schubertová, S.; Krepsová, Z.; Janotková, L.; Potoˇcˇnáková,M.; Kreps, F. -

Trail and Fell Running
Trail and Fell Running in the Yorkshire Dales Trail and Fell Running in the About the Author Pete Ellwood is an experienced fell runner, running and racing in the hills for over thirty years. He has lived Yorkshire Dales in the north all his life and, since his teenage years, has attempted to spend every possible waking hour on the hills and mountains. Pete holds a long service award for volunteering as a member of a mountain rescue team, 40 runs and he completed the Munros in 2003. He regularly com- petes in fell races and mountain marathons in the North in the National Park, of England and Scotland, winning a class of the Saunders Lakeland Mountain Marathon in 2013. including the Three Peaks Pete lives on the edge of the Yorkshire Dales with his wife, two boys and a daily view of Ingleborough to keep by Pete Ellwood him company. Juniper House, Murley Moss, Oxenholme Road, Kendal, Cumbria LA9 7RL www.cicerone.co.uk © Pete Ellwood 2019 Register your book: To sign up to receive free updates, special offers First edition 2019 and GPX files where available, register Contents ISBN: 978 1 85284 922 1 your book at www.cicerone.co.uk. Map key ................................................................7 Printed by KHL Printing, Singapore Acknowledgements Overview map ..........................................................8 A catalogue record for this book is Route summary table ....................................................9 This book would not have been possible available from the British Library. without the help and support of a large ULTRA ................................................................15 © Crown copyright 2019 number of people. I would like to take OS PU100012932 this opportunity to thank all the Settle Harriers who checked routes and posed The Yorkshire Dales ....................................................... -
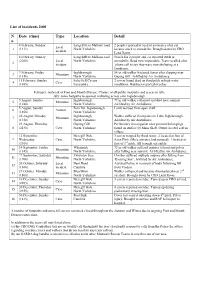
List of Incidents 2001 N O. Date (Time) Type Location Detail
List of incidents 2001 N Date (time) Type Location Detail o. 4 February, Sunday Langcliffe to Malham road 2 people reported in need of assistance after car Local 1 (1311) North Yorkshire became stuck in snowdrifts. Brought down by CRO incident Land Rover. 4 February, Sunday Langcliffe to Malham road Search for 2 people and car reported stuck in (2200) Local North Yorkshire snowdrifts. Road now impassable. Team recalled after 2 incident ‘phone call to say they were now sheltering at a farmhouse. 9 February, Friday Ingleborough 54 yr old walker fractured femur after slipping near 3 Mountain (1150) North Yorkshire Gaping Gill. Airlifted by Air Ambulance. 11 February, Sunday Ireby Fell Cavern 2 cavers found dead on third pitch in high water 4 Cave (1045) Lancashire conditions. Had been reported overdue. February: outbreak of Foot and Mouth Disease. Closure of all public footpaths and access to fells. July: some footpaths re-opened, including access onto Ingleborough. 5 August, Sunday Ingleborough 79 yr old walker collapsed and died near summit. 5 Mountain (1240) North Yorkshire Airlifted by Air Ambulance. 5 August, Sunday Fern Pot, Ingleborough Lamb rescued from open shaft. 6 Animal (1850) North Yorkshire 20 August, Monday Ingleborough Walker suffered chest pains on Little Ingleborough. 7 Mountain (1358) North Yorkshire Airlifted by Air Ambulance. 23 August, Thursday Gaping Gill Preliminary investigation after personal belongings 8 (2130) Cave North Yorkshire found on surface by Main Shaft. Owner located safe in village. 12 September, Meregill Hole 3 cavers trapped by flood water. 2 located at foot of 9 Wednesday Cave North Yorkshire Aven Pitch (Mere entrance had sumped). -

Horton-In- Ribblesdale PEN-Y-GHENT Ribblehead
70 Deepdale 80 686 4. RIBBLEHEAD. 10.4 miles; 5:15 hrs N THREECrag HillPEAKS CHALLENGE Take road NW from Station Inn to ROUTE pass Bleaalongside Viaduct on path for 24 miles (38.6 km) WhernsideMoor . Cross railway line by Cumulative distances and guidance aqueduct and follow path steeply NW 3. HIGH BIRKWITH. 7.0 miles; 3:45 hrs times are shown at each stage. for Dent Dale. Cross fence stile on left Cross road & over small hill to drop to a gate. Timings and distances based on the Continue NW to cross God’s Bridge and on to Whitber Hill route and follow path to Whernside summit Oughtershaw 736 Nether Lodge. Follow farm access road out to WHERNSIDE B6479 and turn right on road to Ribblehead Beckermonds Cam BLACK DUBB MOSS ROUTE WHITBER HILL ROUTE 80 5. WHERNSIDE. 14.2 miles; 7:35 hrs Fell [Until 2013] Go straight on Gearstones [From 2013] Continue Continue S descending gently along ridge at sharp left bend, turning following PW down to the with wall on right, until path bears left NW to cross Hull Pot Beck gate at Horton Scar Lane. steeply downhill to Bruntscar. Follow access at stepping stones. Track Ribblehead Pass this & climb up over road south for The Hill Inn to Philpin Lane, becomes very boggy at P onto Low Sleights Rd. Left to Hill Inn ck e Whitber Hill, bearing left at e n Black Dubb Moss. Go on NW B n i wall corner and continue NW m n a e to cross PW at a stile. -

SLI Chemicals Gmbh Frankfurt Seabuckthorn Pulp Oil Organic Certified
SLI Chemicals GmbH Frankfurt Seabuckthorn Pulp Oil Organic certified The mongolian heart blood of the Emperor – The lemon of the north 15.05.2012 General - The seabuckthorn shrub is native in Central Asia, in the Altai region, Mongolia and China. Seabuckthorn is also growing at European coasts from the Baltic Sea to the Mediterranean Sea. - Seabuckthorn oils play an important role in the Asian and East European medicine. They are often found in pharmaceuticals and cosmetics. - Since ages Seabuckthorn is used in the traditional medicine of Tibet, Mongolia, China and Russia. General - The Latin name is hippophae rhamnoides (hippos for horse). - Seabuckthorn is a pioneer plant and grows also onto extremly poor soil. Seabuckthorn lives in symbiosis with a special microorganism which is able to collect nitrogen from the air and use it like a natural fertilizer. Therefore seabuckthorn improves the soil for other plants. - The plant is growing even in extreme conditions: heat, frost and aridity. - This may be also a reason for the unique composition of the yellow to deep orange berries. - The dioecism of seabuckthorn makes it necessary to have at least a male and a female plant to harvest berries. Seabuckthorn Pulp Oil is dark red and has a fruity aroma and taste. 1 15.05.2012 What is special with the SANDDORN GbR quality aim: get seabuckthorn oil in a native organic quality • There are for methods or process Seabuckthorn Pulp Oil: – A))yg drying the berries to p rocess Seabuckthorn Pulp Oil – B) oil extraction using heat – C) extraction of the pulp oil using hydocarbons, e.g. -

The Ecosystem Effects of Sand-Binding Shrub Hippophae
land Article The Ecosystem Effects of Sand-Binding Shrub Hippophae rhamnoides in Alpine Semi-Arid Desert in the Northeastern Qinghai–Tibet Plateau Lihui Tian 1,2,*, Wangyang Wu 3, Xin Zhou 1, Dengshan Zhang 2, Yang Yu 4, Haijiao Wang 1 and Qiaoyu Wang 1 1 State Key Laboratory of Plateau Ecology and Agriculture, Qinghai University, Xi’ning 810016, China; [email protected] (X.Z.); [email protected] (H.W.); [email protected] (Q.W.) 2 Qinghai Academy of Agricultural Forestry Sciences, Qinghai University, Xi’ning 810016, China; [email protected] 3 School of Earth Sciences, East China University of Technology, Nanchang 200237, China; [email protected] 4 Department of Sediment Research, China Institute of Water Resources and Hydropower Research, Beijing 100038, China; [email protected] * Correspondence: [email protected] Received: 6 November 2019; Accepted: 27 November 2019; Published: 29 November 2019 Abstract: The planting of sand-binding vegetation in the Qinghai Lake watershed at the northeastern edge of the Qinghai–Tibet Plateau began in 1980. For this paper, we took the desert on the eastern shore of Qinghai Lake as the study area. We analyzed a variety of aged Hippophae rhamnoides communities and aeolian activities, and we discuss the relationship between them. The main conclusions are as follows: (1) With an increasing number of binding years, the species composition became more abundant, natural vegetation began to recover, and biodiversity increased year by year. At the same time, plant height, canopy width, and community coverage increased, but H. rhamnoides coverage was reduced to 36.70% as coverage of Artemisia desertorum increased to 25.67% after 10 years of fixing. -

The Pendle Panorama
© Mark Sutcliffe THE PENDLE PANORAMA To the south, the former mill towns of You can see for miles from the top of Pendle, here is a list of mountains visible on a clear day: The prospect of admiring breath-taking views Nelson, Colne, Burnley and Blackburn nestle Clockwise from North: from the summit is what inspires thousands in the valley floor with the fells of the West of visitors to climb Pendle every year. Pennines beyond and on the southwest • Ingleborough (21 miles) • Bleaklow (33) • Parlick Pike (13) Why not provide this information for your horizon, the mountains of Snowdonia. Looking • Whernside (25) • Kinder Scout (38) • Fair Snape (13) visitors via your website or social media? west, the Irish Sea sparkles on the horizon • Pen y Gent (20) • Shining Tor (44) • Hawthornthwaite Fell (15) with Blackpool Tower clearly visible on the • Fountains Fell (19) • Winter Hill (19) • Black Combe (50) To the north, the mountains of the Lake District Fylde Coast. Few places in England offer the can just be glimpsed peaking over the Bowland potential to enjoy such extensive panoramas • Buckden Pike (25) • Clwydian Hills (63) • Ward Stone (17) Fells on the far side of the Ribble Valley. within half an hour of leaving the office. • Great Whernside (24) • Moel Siabod (87) • Wolfhole Crag (15) To the northeast, following the line of the • Thorpe Fell (17) • Snowdon (92) • Scafell Pike (55) Ribble upstream, the views encompass the • Simon’s Seat (21) • Carnedd Llewellyn (85) • Helvellyn (54) famous Three Peaks of the Yorkshire Dales: Ingleborough, Pen-y-ghent and Whernside. -

Wessex-Cave-Club-Journal-Number-216.Pdf
Vol.19 No.216 WESSEX CAVE CLUB JOURNAL March 1988 CONTENTS Editorial 145 Club & Regional Notes 146 Growth At The Grove (Portland) N. Graham 147 Grove Cliff Caves Survey 149 AUSTRALIAN DIARY: CAVING IN NULLARBOR. Pt. 2 (Concluded) N. Hawkes. 151 Christmas in the Wessex (Logbook compilation) 154 BRITISH SPELEOLOGICAL EXPEDITION, SW GUIZHOU, CHINA (Objectives and Aims) P. Weston 156 50 Years Ago 157 From The Log 159 WCC ACCOUNTS 86/87 166 Back Page: Meets etc. Dates, Riddle. NEXT EDITION THE CAVES OF PERIGORD (FRANCE) Pt 1 R. Taviner RAILWAYS FOR CAVE DIGS N. Graham OFFICERS & COMMITTEE OF THE WESSEX CAVE CLUB 1987/88 PRESIDENT LUKE DEVENISH EDITOR NIGEL GRAHAM VICE PRESIDENTS REV. C. CULLINGFORD 60 Williams Ave., Wyke Regis, SIR JOHN WEDGWOOD Weymouth, Dorset DT4 9BP MR P. DOLPHIN Tel: (0305) 789770 (home) HON. CHAIRMAN JIM MOON SALES OFFICER PETE HANN 49 Sommerville Rd., St. Andrews, 3 Queens Terrace, Sherborne, Bristol BS7 9AD Dorset DT9 4ED Tel: (0272) 425391 (home) HUT ADMIN. OFFICER MAIRI RANDS HON. SECRETARY ROB TAVINER c/o Upper Pitts 91 Bristol Road, Whitchurch, COMMITTEE JIM RANDS Bristol BS14 0PS MEMBERS DAVE MORRISON HON. TREASURER BRIAN HANSFORD MALCOLM FOYLE 34 Fleming Rd, Weeke Estate, c/o Upper Pitts Winchester, Hants SO22 6EE LIBRARIANS PETE & ALISON MOODY ASST. SECRETARY MIKE DEWDNEY-YORK NORTHERN CAVING KEITH SANDERSON 23 Northumberland Rd., SECRETARY Heather View, Newby, Redland, Bristol BS6 7AZ Nr. Clapham, Lancs. CAVING SECRETARY PETE WATTS MRO CO-ORDINATOR PETE LACEY c/o Wessex Cave Club 23 Old Banwell Rd., GEAR CURATOR DUNCAN FREW Weston-s-Mare, Avon Brackens, Castledon Rd., Downham, Billericay, Essex CM11 1LD NICK POLLARD HUT WARDENS 32 Holsom Close, Stockwood, Bristol BS14 8LX GRAHAM JOHNSON 5 Dodd Avenue, Wells, Somerset BA4 3TU Opinions expressed in this Journal are not necessarily those of the Editor or of the Wessex Cave as a whole, unless otherwise stated. -

Chemical Composition and Antioxidant Activity of Leaves of Mycorrhized
Chemical composition and antioxidant activity of leaves of mycorrhized sea- buckthorn (Hippophae rhamnoides L.) Anna Jaroszewska1, and Wioletta Biel2* ABSTRACT INTRODUCTION Natural products, either as extracts or as pure compounds, The leaves of sea-buckthorn (Hippophae rhamnoides L.) give limitless possibilities for the discovery of new drugs and can be a rich source of nutrients and biologically active improving food quality, also thanks to their wide availability. substances. Their levels depend on growing conditions, Sea-buckthorn (Hippophae rhamnoides L.) contains a series of agricultural technology and climate. The studies however, compounds including, among others, carotenoids, tocopherols, mainly focus on nutritious value of fruit and seeds and sterols, flavonoids, lipids, ascorbic acid, and tannins. These there is shortage of the information regarding to buckthorn compounds are of interest not only from the chemical point leaves. The aim of the experiment was to determine the of view, but also because many of them exhibit biological and effect of symbiotic mycorrhizal fungi on the chemical therapeutic activities, including antioxidant, antitumor, hepato- composition and antioxidant activity of sea-buckthorn protective, and immunomodulatory properties (Saikia and leaves. The study was conducted in 2014 and 2015 at the Handique, 2013). Evaluation of the nutritional value of sea- Experimental Station in Lipnik, Poland. Mycorrhization buckthorn usually concerns its fruit and seeds, while the leaves improved the nutritional value of leaves of sea-buckthorn are rather neglected. However, some studies confirm that also its by increasing levels of total protein (3%), N free extract leaves are a rich source of compounds with anti-inflammatory (1%), Ca (9%), Na (16%), Fe (18%), Cr (34%), and (Padwad et al., 2006) and antibacterial properties (Upadhyay partially elevating antioxidant activity by increasing the et al., 2011).