Insect Morphology - Legs 1
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
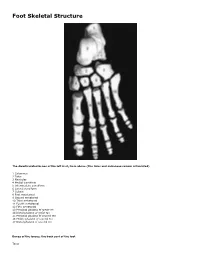
Skeletal Foot Structure
Foot Skeletal Structure The disarticulated bones of the left foot, from above (The talus and calcaneus remain articulated) 1 Calcaneus 2 Talus 3 Navicular 4 Medial cuneiform 5 Intermediate cuneiform 6 Lateral cuneiform 7 Cuboid 8 First metatarsal 9 Second metatarsal 10 Third metatarsal 11 Fourth metatarsal 12 Fifth metatarsal 13 Proximal phalanx of great toe 14 Distal phalanx of great toe 15 Proximal phalanx of second toe 16 Middle phalanx of second toe 17 Distal phalanx of second toe Bones of the tarsus, the back part of the foot Talus Calcaneus Navicular bone Cuboid bone Medial, intermediate and lateral cuneiform bones Bones of the metatarsus, the forepart of the foot First to fifth metatarsal bones (numbered from the medial side) Bones of the toes or digits Phalanges -- a proximal and a distal phalanx for the great toe; proximal, middle and distal phalanges for the second to fifth toes Sesamoid bones Two always present in the tendons of flexor hallucis brevis Origin and meaning of some terms associated with the foot Tibia: Latin for a flute or pipe; the shin bone has a fanciful resemblance to this wind instrument. Fibula: Latin for a pin or skewer; the long thin bone of the leg. Adjective fibular or peroneal, which is from the Greek for pin. Tarsus: Greek for a wicker frame; the basic framework for the back of the foot. Metatarsus: Greek for beyond the tarsus; the forepart of the foot. Talus (astragalus): Latin (Greek) for one of a set of dice; viewed from above the main part of the talus has a rather square appearance. -

Order Ephemeroptera
Glossary 1. Abdomen: the third main division of the body; behind the head and thorax 2. Accessory flagellum: a small fingerlike projection or sub-antenna of the antenna, especially of amphipods 3. Anterior: in front; before 4. Apical: near or pertaining to the end of any structure, part of the structure that is farthest from the body; distal 5. Apicolateral: located apical and to the side 6. Basal: pertaining to the end of any structure that is nearest to the body; proximal 7. Bilobed: divided into two rounded parts (lobes) 8. Calcareous: resembling chalk or bone in texture; containing calcium 9. Carapace: the hardened part of some arthropods that spreads like a shield over several segments of the head and thorax 10. Carinae: elevated ridges or keels, often on a shell or exoskeleton 11. Caudal filament: threadlike projection at the end of the abdomen; like a tail 12. Cercus (pl. cerci): a paired appendage of the last abdominal segment 13. Concentric: a growth pattern on the opercula of some gastropods, marked by a series of circles that lie entirely within each other; compare multi-spiral and pauci-spiral 14. Corneus: resembling horn in texture, slightly hardened but still pliable 15. Coxa: the basal segment of an arthropod leg 16. Creeping welt: a slightly raised, often darkened structure on dipteran larvae 17. Crochet: a small hook-like organ 18. Cupule: a cup shaped organ, as on the antennae of some beetles (Coleoptera) 19. Detritus: disintegrated or broken up mineral or organic material 20. Dextral: the curvature of a gastropod shell where the opening is visible on the right when the spire is pointed up 21. -

Gene Duplication and the Origins Of
Rivera et al. BMC Evolutionary Biology 2010, 10:123 http://www.biomedcentral.com/1471-2148/10/123 Daphnia Genomics Consortium RESEARCH ARTICLE Open Access Gene duplication and the origins of morphological complexity in pancrustacean eyes, a genomic approach Ajna S Rivera1, M Sabrina Pankey1, David C Plachetzki1, Carlos Villacorta1, Anna E Syme1, Jeanne M Serb1,3, Angela R Omilian2, Todd H Oakley1* Abstract Background: Duplication and divergence of genes and genetic networks is hypothesized to be a major driver of the evolution of complexity and novel features. Here, we examine the history of genes and genetic networks in the context of eye evolution by using new approaches to understand patterns of gene duplication during the evolution of metazoan genomes. We hypothesize that 1) genes involved in eye development and phototransduction have duplicated and are retained at higher rates in animal clades that possess more distinct types of optical design; and 2) genes with functional relationships were duplicated and lost together, thereby preserving genetic networks. To test these hypotheses, we examine the rates and patterns of gene duplication and loss evident in 19 metazoan genomes, including that of Daphnia pulex - the first completely sequenced crustacean genome. This is of particular interest because the pancrustaceans (hexapods+crustaceans) have more optical designs than any other major clade of animals, allowing us to test specifically whether the high amount of disparity in pancrustacean eyes is correlated with a higher rate of duplication and retention of vision genes. Results: Using protein predictions from 19 metazoan whole-genome projects, we found all members of 23 gene families known to be involved in eye development or phototransduction and deduced their phylogenetic relationships. -

Canine Tarsus Stabilization Surgical Technique 1 2
Canine Tarsus Stabilization Surgical Technique 1 2 Place the patient in a dorsal recumbent position and Make a medial incision beginning just proximal to the administer general anesthesia. Perform a hanging limb medial malleolus extending distally to the level of the technique with aseptic preparation and appropriate proximal intertarsal joint. Inspect the medial structures limb draping. of the tarsal joint. Identify the origin of the long part of the medial collateral ligament (MCL) on the medial malleolus of the tibia. 3 4 Using the aiming guide, place the 0.049 inch (1.2 mm) Use the 2.0 mm cannulated drill over the guidewire to guidewire from the origin of the long part of the MCL create the tibial bone tunnel for suture passage. in a proximal and slight cranial direction such that the it emerges on the lateral tibia just cranial to the fibula. Make a small incision on the lateral side where the guidewire tents the skin. 5 As a surgical landmark, identify the talocentral tarsal joint on the medial side by either a palpation or by inserting a small needle into the joint space. Using the aiming guide, place a 0.049 inch (1.2 mm) guidewire starting at the medial aspect of the talar head oriented parallel to the talocentral tarsal joint in a slightly plantar direction to capture the calcaneus. The guidewire should emerge just plantar to the boney tubercle of the lateral portion of the distal calcaneus. Make a small incision where the guidewire tents the skin. 6 7 Use the 2.0 mm cannulated drill over the guidewire to Identify the insertion of the short part of the MCL on the create the distal talus bone tunnel for suture passage. -

Equine MRI, Edited by Rachel C
Chapter 17 The p roximal t arsal r egion Rachel Murray , Natasha Werpy , Fabrice Audigi é , Jean - Marie Denoix , Matthew Brokken and Thorben Schulze INTRODUCTION Imaging of the proximal tarsus can be achieved in both standing and anaes- thetized horses. The height of the horse and the amount of hind end mus- culature determines which magnet confi guration will produce the best images of this region. A horse with short limbs and wide, well - muscled quarters may be more diffi cult to fi t into the long, narrow bore of some high - fi eld magnets under general anaesthesia. In the same way, the side of the lateral recumbency should be considered because it is generally easier to bring the proximal tarsus of the lower hind limb closer to the isocentre of the magnet compared with the upper one. Well developed hind limb musculature can prevent the limb from being placed far enough into the bore to allow the tarsus to be positioned at the isocentre of the magnet. The tarsus of a horse with this body type (or stature) is more easily positioned in a standing system. In contrast, for a narrow horse with long limbs, the height of the tarsus and width between the limbs may make it more diffi cult to image this region within a standing system. The degree of sway in the proximal aspect of the limbs in a standing, sedated horse is often substan- tially greater than in the distal aspect of the limbs. This degree of sway can produce signifi cant motion artefact and can make it frustrating to acquire images of the proximal tarsus. -

Periprosthetic Fractures
Periprosthetic Fractures SRS 2017 Stephen R Smith Orthopaedic Surgeon Northeast Nebraska Orthopaedics P C Norfolk Nebraska SRS 2017 Periprosthetic Fractures Fractures around Joint Replacements Mostly Lower Limb Knee Arthroplasty 700,000/ yr. Hip Arthroplasty 350,000/yr. Shoulder Arthroplasty ? 60,000/yr. Elbow Arthroplasty ? 20,000/yr. Periprosthetic Fractures Incidence Increasing due Increasing Demand and High Demands of Older Patients Projections 2025 2,000,000 (2 million) Knee Replacements 750,000 Total Hip Replacements Periprosthetic Fractures Risk Factors Mechanical Patient Factors Implant Loosening Rheumatoid Arthritis Chronic Steroid Use Neurologic Osteolysis Disease/Disorders Osteoporosis Osteopenia Femoral Notching Female Gender (Above TKA) Increasing Age SRS 2017 Periprosthetic Fractures Incidence Hip Intraoperative Acetabulum Cemented 0.2% Uncemented 0.4% During Impaction Under reaming> 2mm, Osteoporosis, Dysplasia Radiation Periprosthetic Fractures Incidence Hip Intraoperative Primary 0.1-5% Classification Osteoporosis,Cementless, Technique, Revision, Minimally Invasive, Revision 3-21% Periprosthetic Fractures Risk Factors DON’T FALL Remove Loose Rugs Minimize Stair Use Rail Stay Home in Bad Weather!!! Use Common Sense SRS 2017 Periprosthetic Fractures Risk Factors This Is Ice DON’T FALL Remove Loose Rugs Minimize Stair Use Rail Stay Home in Bad Weather!!! Use Common Sense SRS 2017 Periprosthetic Fractures Incidence Knee Intraoperative ?? Occasional Medial Femoral Condyle Often Tibial Crack after Stem Impaction Postoperative -

Fracture of the Lesser Trochanter As a Sign of Undiagnosed Tumor Disease in Adults Christian Herren*, Christian D
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by Springer - Publisher Connector Herren et al. Eur J Med Res (2015) 20:72 DOI 10.1186/s40001-015-0167-8 CASE REPORT Open Access Fracture of the lesser trochanter as a sign of undiagnosed tumor disease in adults Christian Herren*, Christian D. Weber, Miguel Pishnamaz, Thomas Dienstknecht, Philipp Kobbe, Frank Hildebrand and Hans‑Christoph Pape Abstract Isolated avulsion fractures of the pelvic ring are rare and occur predominantly in adolescent athletes. Isolated fractures of the lesser trochanter are reported to be pathognomic for tumor diseases in adults. We present a case of a female patient with an isolated avulsion of the lesser trochanter after treatment by her chiropractor. After staging exami‑ nation, we determine the diagnosis of a left-sided carcinoma of the mamma. Additional imaging shows multiple metastases in liver, spine and pelvis. Palliative therapy has started over the course of time. We suggest, on suspicion of a malignant metastatic process, further investigation. Keywords: Fracture, Lesser trochanter, Metastatic, Tumor disease Background described unexplained weight loss of 5 kg in 4 months. Isolated fractures of the lesser trochanter are uncommon Sporadic onset of night sweats was also reported. She had and have been reported predominantly in adolescent ath- no other musculoskeletal or constitutional diseases in her letes [1]. This injury is caused by severe impact, usually medical history. Physical examination showed tenderness in context of contact sports and following a forceful and in the right groin, almost preserved passive mobility of sudden muscle contraction of the iliopsoas with avulsion the right hip joint in the full range of motion. -
Supplementary Table 1: Description of All Clinical Tests Test Protocol
Supplementary Table 1: Description of all clinical tests Test Protocol description Tibiofemoral • Palpate & mark tibial tuberosity & midpoint over the talus neck frontal plane • Ask participant to stand on footprint map with foot at 10° external rotation, feet shoulder width, looking alignment forward, 50% weightbearing • Place callipers of inclinometer in alignment with the the two landmarks • Record varus/valgus direction in degrees Herrington test • Participant supine on plinth, knee positioned and supported in 20° of knee flexion (to place the patella within the trochlea groove) • With knee in position, place a piece of 1” Leukotape (or similar) across the knee joint, and mark the medial and lateral epicondyles of the femur and mid-point of the patella. Be sure to make note of medial and lateral end of tape • Repeat 3 times, attaching tape to this document for measuring later 30 second chair • Shoes on, middle of chair, feet ~ shoulder width apart and slightly behind knees with feet flat on floor, stand test arms crossed on chest • Instructions “stand up keeping arms across chest, and ensure you stand completely up so hips and knees are fully extended; then sit completely back down, so that the bottom fully touches the seat, as many times as possible in 30 seconds,” • 1-2 practice repetitions for technique • One 30-second test trial • Record number of correctly performed full stands (if more than ½ of way up at end of the test, counted as a full stand) Repetitive single • Shoes on, seated on edge of plinth, foot placed with heel 10 cm forward from a plumb line at edge of leg rise test plinth, other leg held at side of body, arms across chest. -

The Evolution and Development of Arthropod Appendages
Evolving Form and Function: Fossils and Development Proceedings of a symposium honoring Adolf Seilacher for his contributions to paleontology, in celebration of his 80th birthday Derek E. G. Briggs, Editor April 1– 2, 2005 New Haven, Connecticut A Special Publication of the Peabody Museum of Natural History Yale University New Haven, Connecticut, U.S.A. December 2005 Evolving Form and Function: Fossils and Development Proceedings of a symposium honoring Adolf Seilacher for his contributions to paleontology, in celebration of his 80th birthday A Special Publication of the Peabody Museum of Natural History, Yale University Derek E.G. Briggs, Editor These papers are the proceedings of Evolving Form and Function: Fossils and Development, a symposium held on April 1–2, 2005, at Yale University. Yale Peabody Museum Publications Jacques Gauthier, Curatorial Editor-in-Chief Lawrence F. Gall, Executive Editor Rosemary Volpe, Publications Editor Joyce Gherlone, Publications Assistant Design by Rosemary Volpe • Index by Aardvark Indexing Cover: Fossil specimen of Scyphocrinites sp., Upper Silurian, Morocco (YPM 202267). Purchased for the Yale Peabody Museum by Dr. Seilacher. Photograph by Jerry Domian. © 2005 Peabody Museum of Natural History, Yale University. All rights reserved. Frontispiece: Photograph of Dr. Adolf Seilacher by Wolfgang Gerber. Used with permission. All rights reserved. In addition to occasional Special Publications, the Yale Peabody Museum publishes the Bulletin of the Peabody Museum of Natural History, Postilla and the Yale University Publications in Anthropology. A com- plete list of titles, along with submission guidelines for contributors, can be obtained from the Yale Peabody Museum website or requested from the Publications Office at the address below. -

The Occurrence of Tarsal Injuries in Male Mice of C57BL/6N Substrains in Multiple
bioRxiv preprint doi: https://doi.org/10.1101/2020.02.25.964254; this version posted February 25, 2020. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available under aCC-BY 4.0 International license. 1 Long title: The occurrence of tarsal injuries in male mice of C57BL/6N substrains in multiple 2 international mouse facilities 3 4 Short title: Tarsal injury in C57BL/6N male mice 5 6 Eleanor Herbert2, Michelle Stewart1, Marie Hutchison1, Ann M. Flenniken4,5, Dawei Qu4,5, 7 Lauryl M. J. Nutter4,6, Colin McKerlie4,6, Liane Hobson1, Brenda Kick3, Bonnie Lyons3, Jean- 8 Paul Wiegand3, Rosalinda Doty3, Juan Antonio Aguilar-Pimentel7, Martin Hrabe de Angelis7,8,9, 9 Mary Dickinson10, John Seavitt10, Jacqueline K. White3, Cheryl L Scudamore1, Sara Wells1* 10 11 12 13 Affiliations 14 1 Mary Lyon Centre, MRC Harwell Institute, Harwell Campus, Oxfordshire, OX11 0RD, UK 15 2 Department of Pathobiology and Population Sciences, Royal Veterinary College, Hertfordshire, 16 UK AL9 7TA, UK 17 3 The Jackson Laboratory, 600 Main Street, Bar Harbor, Maine 04609, USA 18 4 The Centre for Phenogenomics, Toronto, ON, Canada, M5T 3H7 19 5 Lunenfeld-Tanenbaum Research Institute, Sinai Health, 600 University Avenue, Toronto, 20 ON, Canada, M5G 1X5 21 6 The Hospital for Sick Children, 555 University Avenue, Toronto, ON, Canada, M5G 1X8 22 7German Mouse Clinic, Institute of Experimental Genetics, Helmholtz Zentrum München, 23 Ingolstädter Landstraße 1, 85764 Neuherberg, Germany 24 8School of Life Science Weihenstephan, Technische Universität München, Alte Akademie 8, 25 85354 Freising, Germany 26 9German Center for Diabetes Research (DZD), Ingolstädter Landstr. -

Exploring the Origin of Insect Wings from an Evo-Devo Perspective
Available online at www.sciencedirect.com ScienceDirect Exploring the origin of insect wings from an evo-devo perspective Courtney M Clark-Hachtel and Yoshinori Tomoyasu Although insect wings are often used as an example of once (i.e. are monophyletic) sometime during the Upper morphological novelty, the origin of insect wings remains a Devonian or Lower Carboniferous (370-330 MYA) [3,5,9]. mystery and is regarded as a major conundrum in biology. Over By the early Permian (300 MYA), winged insects had a century of debates and observations have culminated in two diversified into at least 10 orders [4]. Therefore, there is prominent hypotheses on the origin of insect wings: the tergal quite a large gap in the fossil record between apterygote hypothesis and the pleural hypothesis. However, despite and diverged pterygote lineages, which has resulted in a accumulating efforts to unveil the origin of insect wings, neither long-running debate over where insect wings have come hypothesis has been able to surpass the other. Recent from and how they have evolved. investigations using the evolutionary developmental biology (evo-devo) approach have started shedding new light on this Two proposed wing origins century-long debate. Here, we review these evo-devo studies The insect wing origin debate can be broken into two and discuss how their findings may support a dual origin of main groups of thought; wings evolved from the tergum of insect wings, which could unify the two major hypotheses. ancestral insects or wings evolved from pleuron-associat- Address ed structures (Figure 1. See Box 1 for insect anatomy) Miami University, Pearson Hall, 700E High Street, Oxford, OH 45056, [10 ]. -

Radiography and Radiographic Interpretation of the Tarsus (VET-473) Western Veterinary Conference 2004 Timothy R
Radiography and Radiographic Interpretation of the Tarsus (VET-473) Western Veterinary Conference 2004 Timothy R. O'Brien, DVM, PhD, DACVR University of California, Davis Davis, CA, USA 18276161 I. Introduction Tarsal diseases are a common cause of hind limb lameness in the horse. The clinical signs of tarsal disease often resemble those of stifle disease particularly the response to flexion tests. As a result both the stifle and tarsus are commonly radiographed to identify pathologic change in the horse with a hind limb lameness and during purchase evaluations. It seems prudent for purchase evaluations to radiograph these anatomical areas in both limbs. Radiographic interpretation of the tarsus is considered difficult by most equine practitioners. There seems to be three major causes of this difficulty. These causes relate to the numerous bones comprising the tarsus, variety of shapes of the tarsal bones, and veterinarians tend to be less familiar with the radiographic anatomy and pathology of the tarsus than most other anatomical regions, particularly regions of the forelimb. I am going to make some general statements in an attempt to simplify the radiographic interpretation of the tarsus. These general statements are: Most radiographic findings are located at the dorsum of the tarsus. Pathologic abnormalities of the tarsus tend to be in specific anatomical areas. Soft tissue swelling (STS) is a key for assisting to localize the abnormality. There are only a few radiographic diagnoses for the tarsus. The diagnoses of abnormalities of the tarsus identified with radiology can be classified in the following way: 1. Soft tissue swelling without bony change 2.