Perspectives
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

An Eloreta Quantitative EEG Study
brain sciences Article Power Spectral Differences between Transient Epileptic and Global Amnesia: An eLORETA Quantitative EEG Study Jacopo Lanzone 1,* , Claudio Imperatori 2 , Giovanni Assenza 1 , Lorenzo Ricci 1 , Benedetto Farina 2, Vincenzo Di Lazzaro 1 and Mario Tombini 1 1 Neurology, Neurophysiology and Neurobiology Unit, Department of Medicine, Università Campus Bio-Medico di Roma, 00128 Rome, Italy; [email protected] (G.A.); [email protected] (L.R.); [email protected] (V.D.L.); [email protected] (M.T.) 2 Cognitive and Clinical Psychology Laboratory, Department of Human Sciences, European University of Rome, Via degli Aldobrandeschi 190, 00163 Rome, Italy; [email protected] (C.I.); [email protected] (B.F.) * Correspondence: [email protected] Received: 14 July 2020; Accepted: 4 September 2020; Published: 6 September 2020 Abstract: Transient epileptic amnesia (TEA) is a rare epileptic condition, often confused with transient global amnesia (TGA). In a real-life scenario, differential diagnosis between these two conditions can be hard. In this study we use power spectral analysis empowered by exact Low Resolution Brain Electromagnetic Tomography (eLORETA) to evidence the differences between TEA and TGA. Fifteen patients affected by TEA (64.2 5.2 y.o.; 11 female/4 male; 10 left and 5 right temporal epileptic focus) ± and 15 patients affected by TGA (65.8 7.2 y.o.; 11 females/4 males) were retrospectively identified in ± our clinical records. All patients recorded EEGs after symptoms offset. EEGs were analyzed with eLORETA to evidence power spectral contrast between the two conditions. We used an inverse problem solution to localize the source of spectral differences. -

Toward a Common Terminology for the Gyri and Sulci of the Human Cerebral Cortex Hans Ten Donkelaar, Nathalie Tzourio-Mazoyer, Jürgen Mai
Toward a Common Terminology for the Gyri and Sulci of the Human Cerebral Cortex Hans ten Donkelaar, Nathalie Tzourio-Mazoyer, Jürgen Mai To cite this version: Hans ten Donkelaar, Nathalie Tzourio-Mazoyer, Jürgen Mai. Toward a Common Terminology for the Gyri and Sulci of the Human Cerebral Cortex. Frontiers in Neuroanatomy, Frontiers, 2018, 12, pp.93. 10.3389/fnana.2018.00093. hal-01929541 HAL Id: hal-01929541 https://hal.archives-ouvertes.fr/hal-01929541 Submitted on 21 Nov 2018 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. REVIEW published: 19 November 2018 doi: 10.3389/fnana.2018.00093 Toward a Common Terminology for the Gyri and Sulci of the Human Cerebral Cortex Hans J. ten Donkelaar 1*†, Nathalie Tzourio-Mazoyer 2† and Jürgen K. Mai 3† 1 Department of Neurology, Donders Center for Medical Neuroscience, Radboud University Medical Center, Nijmegen, Netherlands, 2 IMN Institut des Maladies Neurodégénératives UMR 5293, Université de Bordeaux, Bordeaux, France, 3 Institute for Anatomy, Heinrich Heine University, Düsseldorf, Germany The gyri and sulci of the human brain were defined by pioneers such as Louis-Pierre Gratiolet and Alexander Ecker, and extensified by, among others, Dejerine (1895) and von Economo and Koskinas (1925). -

The Pre/Parasubiculum: a Hippocampal Hub for Scene- Based Cognition? Marshall a Dalton and Eleanor a Maguire
Available online at www.sciencedirect.com ScienceDirect The pre/parasubiculum: a hippocampal hub for scene- based cognition? Marshall A Dalton and Eleanor A Maguire Internal representations of the world in the form of spatially which posits that one function of the hippocampus is to coherent scenes have been linked with cognitive functions construct internal representations of scenes in the ser- including episodic memory, navigation and imagining the vice of memory, navigation, imagination, decision-mak- future. In human neuroimaging studies, a specific hippocampal ing and a host of other functions [11 ]. Recent inves- subregion, the pre/parasubiculum, is consistently engaged tigations have further refined our understanding of during scene-based cognition. Here we review recent evidence hippocampal involvement in scene-based cognition. to consider why this might be the case. We note that the pre/ Specifically, a portion of the anterior medial hippocam- parasubiculum is a primary target of the parieto-medial pus is consistently engaged by tasks involving scenes temporal processing pathway, it receives integrated [11 ], although it is not yet clear why a specific subre- information from foveal and peripheral visual inputs and it is gion of the hippocampus would be preferentially contiguous with the retrosplenial cortex. We discuss why these recruited in this manner. factors might indicate that the pre/parasubiculum has privileged access to holistic representations of the environment Here we review the extant evidence, drawing largely from and could be neuroanatomically determined to preferentially advances in the understanding of visuospatial processing process scenes. pathways. We propose that the anterior medial portion of the hippocampus represents an important hub of an Address extended network that underlies scene-related cognition, Wellcome Trust Centre for Neuroimaging, Institute of Neurology, and we generate specific hypotheses concerning the University College London, 12 Queen Square, London WC1N 3BG, UK functional contributions of hippocampal subfields. -

A. HM, B. Spa@Al Naviga@On 2. Hippocampus
Outline 1. What does Hippocampus do? A. HM, B. Spaal navigaon 2. Hippocampus anatomy and connec;vity CA1, CA3 diagram etc. Connec;vity to other regions. 3. Place fields in Hippocampus – show examples + the O'Keefe effect 4. Plas;city of place fields + NMDA blocker + Morris water maze 5. Replay of sequences 6. Entorhinal grid cells What does the Hippocampus do? 1. Memory 2. Spaal representaon Henry Gustav Molaison (February 26, 1926 – December 2, 2008), beger known as HM “AYer operaon this young man could no longer recognize the hospital staff nor find his way to the bathroom, and he seemed to recall nothing of the day-to-day events of his hospital life. There was also a par;al retrograde amnesia, inasmuch as he did not remember the death of a favorite uncle three years previously, nor anything of the period in the hospital, yet could recall some trivial events that had occurred just before his admission to the hospital. His early memories were apparently vivid and intact. This paent’s memory defect has persisted without improvement to the present ;me, and numerous illustraons of its severity could be given. Ten months ago the family moved from their old house to a new one a few blocks away on the same street; he s;ll has not learned the new address, though remembering the old one perfectly, nor can he be trusted to find his way home alone.” “Moreover, he does not know where objects in con;nual use are kept; for example, his mother s;ll has to tell him where to find the lawn mower, even though he may have been using it only the day before. -

Retrohippocampal Cortical Neurons During Hippocampal Sharp Waves in the Behaving Rat
The Journal of Neuroscience, October 1994, 74(10): 6160-6170 Selective Activation of Deep Layer (V-VI) Retrohippocampal Cortical Neurons during Hippocampal Sharp Waves in the Behaving Rat J. J. Chrobak and G. Buzsaki Center for Molecular and Behavioral Neuroscience, Rutgers, The State University of New Jersey, Newark, New Jersey 07102 The coordinated activity of hippocampal neurons is reflected feet on their postsynaptic neocortical targets and may rep- by macroscopic patterns, theta and sharp waves (SPW), ev- resent a physiological mechanism for memory trace transfer ident in extracellular field recordings. The importance of these from the hippocampus to the neocortex. patterns is underscored by the ordered relation of specific [Key words: hippocampus, entorhinal cortex, theta, sharp neuronal populations to each pattern as well as the relation waves, oscillations, memory, temporal lobe epilepsy, Ab- of each pattern to distinct behavioral states. During awake heimer’s disease] immobility, consummatory behavior, and slow wave sleep, CA3 and CA1 neurons participate in organized population Retrohippocampal structures [entorhinal cortex (EC), parasub- bursts during SPW. In contrast, during theta-associated ex- iculum, presubiculum, and subiculum] process and transmit ploratory activity, the majority of principle cells are silent. information between the neocortex and the hippocampus. The Considerably less is known about the discharge properties electrophysiology of thesestructures has received scant attention of retrohippocampal neurons during theta, and particularly despite their importance as a substrate for memory (Amaral, during SPW. These retrohippocampal neurons (entorhinal 1987; Zola-Morgan et al., 1989; Squire, 1992)and asfocal point cortical, parasubicular, presubicular, and subicular) process for the pathophysiology of dementia (Hyman et al., 1984; Van and transmit information between the neocortex and the hip- Hoesenet al., 1991) and temporal lobe epilepsy (Rutecki et al., pocampus. -

The Surgical Prognostic Significance of the Electroencephalographic Prediction of Ammon's Horn Sclerosis in Epileptics by W
J Neurol Neurosurg Psychiatry: first published as 10.1136/jnnp.21.1.24 on 1 February 1958. Downloaded from J. Neurol. Neurosurg. Psychiat., 1958, 21, 24. THE SURGICAL PROGNOSTIC SIGNIFICANCE OF THE ELECTROENCEPHALOGRAPHIC PREDICTION OF AMMON'S HORN SCLEROSIS IN EPILEPTICS BY W. A. KENNEDY and DENIS HILL BASED ON PATHOLOGICAL MATERIAL SUPPLIED BY J. B. CAVANAGH and A. MEYER From the Institute ofPsychiatry and the Guy's-Maudsley Neurosurgical Unit, London In a previous communication (Falconer, Hill, Jasper (1954) and recently by Rasmussen (1957). Meyer, Mitchell, and Pond, 1955) the results of The diagnostic and pathological data and the treating intractable temporal lobe epilepsy by tem- therapeutic effects of temporal lobectomy carried poral lobectomy were given. A survey was reported out on a larger series of 50 cases are now available of the clinical, electroencephalographic, radiological, and a preliminary report has been given (Falconer, and pathological findings in 31 cases. The follow-up Meyer, Hill, and Wilson, 1957). Following Earle etguest. Protected by copyright. study of the patients showed beneficial effects upon al. (1953) the pathological findings have been classi- the epilepsy and upon the personality disorder in a fied in two main groups. While all cases in which a proportion which compared very favourably with space-occupying lesion could be anticipated on most published series. The opinion was expressed clinical, radiological, or electroencephalographic that this was due to inclusion of the uncus, Ammon's grounds were excluded from the series, there were, horn, and possibly the amygdaloid nucleus in the nevertheless, 14 cases in which very small focal resected tissue. -

Creating a Web-Based 3D Interactive Resource to Teach the Anatomy of the Human Hippocampus
Morphology of Memory: Creating a web-based 3D interactive resource to teach the anatomy of the human hippocampus by Alisa M. Brandt A thesis submitted to Johns Hopkins University in conformity with the requirements for the degree of Master of Arts Baltimore, Maryland March, 2019 © 2019 Alisa Brandt All Rights Reserved ABSTRACT The hippocampus is a critical region of the brain involved in memory and learning. It has been widely researched in animals and humans due to its role in consolidating new experiences into long-term declarative memories and its vulnerability in neurodegenerative diseases. The hippocampus is a complex, curved structure containing many interconnected regions that consist of distinct cell types. Despite the importance of understanding the normal state of hippocampal anatomy for studying its functions and the disease processes that affect it, didactic educational resources are severely limited. The literature on the hippocampus is expansive and detailed, but a communication gap exists between researchers presenting hippocampal data and those seeking to improve their understanding of this part of the brain. The hippocampus is typically viewed in a two-dimensional fashion; students and scientists have diffculty visualizing its three-dimensional anatomy and its structural relationships in space. To improve understanding of the hippocampus, an interactive, web-based educational resource was created containing a pre-rendered 3D animation and manipulable 3D models of hippocampal regions. Segmentations of magnetic resonance imaging data were modifed and sculpted to build idealized anatomical models suitable for teaching purposes. These models were animated in combination with illustrations and narration to introduce the viewer to the subject, and the completed animation was uploaded online and embedded into the interactive. -

Subtemporal Transparahippocampal Amygdalohippocampectomy for Surgical Treatment of Mesial Temporal Lobe Epilepsy Technical Note
Subtemporal transparahippocampal amygdalohippocampectomy for surgical treatment of mesial temporal lobe epilepsy Technical note T. S. Park, M.D., Blaise F. D. Bourgeois, M.D., Daniel L. Silbergeld, M.D., and W. Edwin Dodson, M.D. Department of Neurology and Neurological Surgery, Washington University School of Medicine, and St. Louis Children's Hospital, St. Louis, Missouri Amygdalohippocampectomy (AH) is an accepted surgical option for treatment of medically refractory mesial temporal lobe epilepsy. Operative approaches to the amygdala and hippocampus that previously have been reported include: the sylvian fissure, the superior temporal sulcus, the middle temporal gyrus, and the fusiform gyrus. Regardless of the approach, AH permits not only extirpation of an epileptogenic focus in the amygdala and anterior hippocampus, but interruption of pathways of seizure spread via the entorhinal cortex and the parahippocampal gyrus. The authors report a modification of a surgical technique for AH via the parahippocampal gyrus, in which excision is limited to the anterior hippocampus, amygdala and parahippocampal gyrus while preserving the fusiform gyrus and the rest of the temporal lobe. Because transparahippocampal AH avoids injury to the fusiform gyrus and the lateral temporal lobe, it can be performed without intracarotid sodium amobarbital testing of language dominance and language mapping. Thus the operation would be particularly suitable for pediatric patients in whom intraoperative language mapping before resection is difficult. Key Words * amygdalohippocampectomy * complex partial seizure * parahippocampal gyrus * subtemporal approach Currently several different variations of temporal lobe resections are used for medically intractable complex partial seizures.[4,6,8,18,21,30,34] Among these operations is amygdalohippocampectomy (AH), first described in 1958 by Niemeyer,[16] who approached the amygdala and hippocampus through an incision on the middle temporal gyrus. -

Traumatic Early Life Stress in the Developing Hippocampus: a Meta-Analysis of MRI Studies
Walden University ScholarWorks Walden Dissertations and Doctoral Studies Walden Dissertations and Doctoral Studies Collection 2020 Traumatic Early Life Stress in the Developing Hippocampus: A Meta-Analysis of MRI Studies Sharon Johnson Walden University Follow this and additional works at: https://scholarworks.waldenu.edu/dissertations Part of the Psychology Commons This Dissertation is brought to you for free and open access by the Walden Dissertations and Doctoral Studies Collection at ScholarWorks. It has been accepted for inclusion in Walden Dissertations and Doctoral Studies by an authorized administrator of ScholarWorks. For more information, please contact [email protected]. Walden University College of Social and Behavioral Sciences This is to certify that the doctoral dissertation by Sharon Lee Johnson has been found to be complete and satisfactory in all respects, and that any and all revisions required by the review committee have been made. Review Committee Dr. Scott Wowra, Committee Chairperson, Psychology Faculty Dr. Patricia Costello, Committee Member, Psychology Faculty Dr. Kimberley Cox, University Reviewer, Psychology Faculty Chief Academic Officer and Provost Sue Subocz, Ph.D. Walden University 2020 Abstract Traumatic Early Life Stress in the Developing Hippocampus: A Meta-Analysis of MRI Studies by Sharon Lee Johnson MPhil, Walden University, 2019 M.Ed., Youngstown State University, 1994 BA, Kent State University, 1979 Dissertation Submitted in Partial Fulfillment of the Requirements for the Degree of Doctor of Philosophy Health Psychology Walden University August 2020 Abstract Advancements in neuroimaging techniques afford researchers the opportunity to examine the actual brains of living persons, which exponentially contributes to new insights regarding brain and behavior phenomena. However, empirical studies investigating stress and the hippocampus attend primarily to adult populations - less on children and adolescents. -

1. Lateral View of Lobes in Left Hemisphere TOPOGRAPHY
TOPOGRAPHY T1 Division of Cerebral Cortex into Lobes 1. Lateral View of Lobes in Left Hemisphere 2. Medial View of Lobes in Right Hemisphere PARIETAL PARIETAL LIMBIC FRONTAL FRONTAL INSULAR: buried OCCIPITAL OCCIPITAL in lateral fissure TEMPORAL TEMPORAL 3. Dorsal View of Lobes 4. Ventral View of Lobes PARIETAL TEMPORAL LIMBIC FRONTAL OCCIPITAL FRONTAL OCCIPITAL Comment: The cerebral lobes are arbitrary divisions of the cerebrum, taking their names, for the most part, from overlying bones. They are not functional subdivisions of the brain, but serve as a reference for locating specific functions within them. The anterior (rostral) end of the frontal lobe is referred to as the frontal pole. Similarly, the anterior end of the temporal lobe is the temporal pole, and the posterior end of the occipital lobe the occipital pole. TOPOGRAPHY T2 central sulcus central sulcus parietal frontal occipital lateral temporal lateral sulcus sulcus SUMMARY CARTOON: LOBES SUMMARY CARTOON: GYRI Lateral View of Left Hemisphere central sulcus postcentral superior parietal superior precentral gyrus gyrus lobule frontal intraparietal sulcus gyrus inferior parietal lobule: supramarginal and angular gyri middle frontal parieto-occipital sulcus gyrus incision for close-up below OP T preoccipital O notch inferior frontal cerebellum gyrus: O-orbital lateral T-triangular sulcus superior, middle and inferior temporal gyri OP-opercular Lateral View of Insula central sulcus cut surface corresponding to incision in above figure insula superior temporal gyrus Comment: Insula (insular gyri) exposed by removal of overlying opercula (“lids” of frontal and parietal cortex). TOPOGRAPHY T3 Language sites and arcuate fasciculus. MRI reconstruction from a volunteer. central sulcus supramarginal site (posterior Wernicke’s) Language sites (squares) approximated from electrical stimulation sites in patients undergoing operations for epilepsy or tumor removal (Ojeman and Berger). -
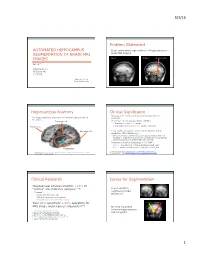
5/3/16 1 Automated Hippocampus Segmentation of Brain
5/3/16 Problem Statement AUTOMATED HIPPOCAMPUS • Goal: automated segmentation of hippocampus in SEGMENTATION OF BRAIN MRI brain MRI images Input Output hippocampus IMAGES Group 7 Yingchuan Hu Chunyan Wu Li Zhong May 1st, 2014 Cornell University Hippocampus Anatomy Clinical Significance • Epilepsy is the most common serious brain disorder The hippocampus is located in the medial temporal lobe of worldwide. the brain. 1 Temporal lobe • Prevalence of epilepsy worldwide (WHO ) • 7 sufferers in every 1,000 people • 3 new sufferers in every 10,000 people each year Frontal lobe Occipital lobe • People with epilepsy are at increased risks for status epilepticus (life-threatening) • One continuous, unremitting seizure lasting longer than five minutes or recurrent seizures without regaining consciousness between seizures for greater than five minutes. • Prevalence of status epilepticus in US (NIH2) • 195,000 new patients of status epilepticus each year • 42,000 deaths caused by status epilepticus each year 1. World Health Organization http://www.who.int/mental_health/neurology/epilepsy/en/ *Modified from a scan of a plate of “Posterior and inferior cornua of left lateral ventricle exposed 2. National institute of Health http://www.ninds.nih.gov/disorders/epilepsy/detail_epilepsy.htm from the side” in Gary’s Anatomy Clinical Research Issues for Segmentation • Hippocampal volume reduction >10% of “normal” size indicates epilepsy.[1-4] • Low contrast to neighboring brain • “normal”: structures • People with the same age • Bilateral hippocampus comparison • Personal changes in more than 1 year • Over 90% sensitivity + 98% specificity for amgydala hippocampus [5-7] MRI image measurement diagnosis. • No clear boundary between hippocampus 1. Cook MJ, et al. -
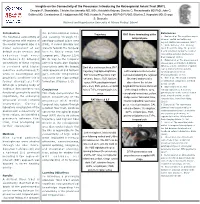
Insights on the Connectivity of the Precuneus: Introducing the Retrosplenial Aslant Tract (RAT)
Insights on the Connectivity of the Precuneus: Introducing the Retrosplenial Aslant Tract (RAT). Georgios P. Skandalakis; Christos Koutsarnakis MD, MSc; Aristotelis Kalyvas; Dimitris G. Placantonakis MD PhD; John G. Golfinos MD; Constantinos G. Hadjipanayis MD PhD; Kostas N. Fountas MD PhD FAANS; Eftychia Z. Kapsalaki MD; George S. Stranjalis National and Kapodistrian University of Athens Medical School Introduction the parieto-occipital sulcus, References Trajectory RAT fibers terminating at the The functional connectivity of and passing through the 1. Epstein et al. The cognitive map in temporal pole humans: spatial navigation and the precuneus with regions of parahippocampal place area beyond. Nature neuroscience. (2017) the medial temporal lobe is a (PPA), it curves laterally and 2. Wade-Bohleber et al. Thinking major component of our projects towards the temporal about the past to shape the present: default mode network and horn to finally reach the Neural activation during the recall of relationship episodes. Behavioural underlies high order temporal pole. (Figures 1,3,4) Brain Research (2018). functions.(1-5) Stronger On its way to the temporal 3. Hebscher et al. The precuneus and connectivity of these regions pole this tracts also displays hippocampus contribute to individual differences in the unfolding of spatial is corelated with higher connections with the lingual, Dark blue continuous lines, RAT representations during episodic cognitive performances(7) parahippocampal and fusiform trajectory; highlighted light blue, Left hemisphere infero-medial autobiographical memory. while in neurological and gyri, inferior longitudinal RAT Cortical Projections; CaF, view demonstrating the regional Neuropsychologia. (2018) psychiatric conditions this is fasciculus and hippocampal calcarine fissure; FuG, fusiform fiber tract anatomy after 4.