Fire in the Southeastern Deserts Bioregion
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Palm Haven Garden Plant List - 2010
PALM HAVEN GARDEN PLANT LIST - 2010 BOTANICAL NAME COMMON NAME Achillea millefolium 'Paprika' Paprika yarrow Arctostaphylos glauca big berry manzanita Arctostaphylos pajaroensis Pajaro manzanita Arctostaphylos uva-ursi bearberry Arctostaphylos uva-ursi 'San Bruno Mountain' San Bruno Mountain bearberry Aristolochia californica California pipevine Artemisia pycnocephala coastal sagewort Baccharis pilularis 'Twin Peaks' Twin Peaks dwarf coyote brush Berberis aquifolium 'Golden Abundance' Golden Abundance Oregon-grape Carex spissa San Diego sedge Ceanothus 'Joyce Coulter' Joyce Coulte wild lilac Ceanothus arboreus feltleaf ceanothus Ceanothus gloriosus var exaltatus 'Emily Brown' Emily Brown glory brush Dendromecon harfordii Channel Island tree poppy Epilobium canum California fuchsia Erigeron glaucus seaside daisy Eriogonum fasciculatum California buckwheat Eriogonum latifolium coast buckwheat Eriogonum umbellatum sulfur flower Eriophyllum confertiflorum golden yarrow Heteromeles arbutifolia toyon Heterotheca sessilifolora false goldenaster Iris douglasiana Douglas iris Leymus condensatus 'Canyon Prince' Canyon Prince giant wild rye Mimulus aurantiacus sticky monkeyflower Monardella villosa coyote mint Mulhenbergia rigens deer grass Penstemon heterophyllus foothill penstemon Physocarpus capitatus ninebark Rhamnus californica 'Ed Holm' Ed Holm coffeeberry Rhamnus californica 'Mound San Bruno' Mound San Bruno coffeeberry Salvia clevelandii Cleveland sage Salvia mellifera 'Shirley's Creeper' Shirley's Creepe sage Salvia sonomensis 'Bee's Bliss' Bee's Bliss sage Satureja douglasii yerba buena Trichostema lanatum wolly blue curls Typha angustifolia narrow-leaved cattail Verbena lilacina lilac verbena Vitis californica 'Roger's Red' Roger's Red California wild grape. -
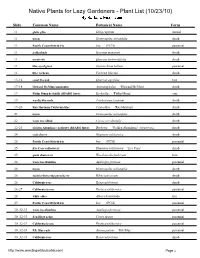
Native Plants for Lazy Gardeners - Plant List (10/23/10)
Native Plants for Lazy Gardeners - Plant List (10/23/10) Slide Common Name Botanical Name Form 11 globe gilia Gilia capitata annual 11 toyon Heteromeles arbutifolia shrub 11 Pacific Coast Hybrid iris Iris (PCH) perennial 11 goldenbush Isocoma menziesii shrub 11 scrub oak Quercus berberidifolia shrub 11 blue-eyed grass Sisyrinchium bellum perennial 11 lilac verbena Verbena lilacina shrub 13-16 coast live oak Quercus agrifolia tree 17-18 Howard McMinn man anita Arctostaphylos 'Howard McMinn' shrub 19 Philip Mun keckiella (RSABG Intro) Keckiella 'Philip Munz' ine 19 woolly bluecurls Trichostema lanatum shrub 19-20 Ray Hartman California lilac Ceanothus 'Ray Hartman' shrub 21 toyon Heteromeles arbutifolia shrub 22 western redbud Cercis occidentalis shrub 22-23 Golden Abundance barberry (RSABG Intro) Berberis 'Golden Abundance' (MAHONIA) shrub 2, coffeeberry Rhamnus californica shrub 25 Pacific Coast Hybrid iris Iris (PCH) perennial 25 Eve Case coffeeberry Rhamnus californica '. e Case' shrub 25 giant chain fern Woodwardia fimbriata fern 26 western columbine Aquilegia formosa perennial 26 toyon Heteromeles arbutifolia shrub 26 fuchsia-flowering gooseberry Ribes speciosum shrub 26 California rose Rosa californica shrub 26-27 California fescue Festuca californica perennial 28 white alder Alnus rhombifolia tree 29 Pacific Coast Hybrid iris Iris (PCH) perennial 30 032-33 western columbine Aquilegia formosa perennial 30 032-33 San Diego sedge Carex spissa perennial 30 032-33 California fescue Festuca californica perennial 30 032-33 Elk Blue rush Juncus patens '.l1 2lue' perennial 30 032-33 California rose Rosa californica shrub http://www weedingwildsuburbia com/ Page 1 30 032-3, toyon Heteromeles arbutifolia shrub 30 032-3, fuchsia-flowering gooseberry Ribes speciosum shrub 30 032-3, Claremont pink-flowering currant (RSA Intro) Ribes sanguineum ar. -

Global Seagrass Distribution and Diversity: a Bioregional Model ⁎ F
Journal of Experimental Marine Biology and Ecology 350 (2007) 3–20 www.elsevier.com/locate/jembe Global seagrass distribution and diversity: A bioregional model ⁎ F. Short a, , T. Carruthers b, W. Dennison b, M. Waycott c a Department of Natural Resources, University of New Hampshire, Jackson Estuarine Laboratory, Durham, NH 03824, USA b Integration and Application Network, University of Maryland Center for Environmental Science, Cambridge, MD 21613, USA c School of Marine and Tropical Biology, James Cook University, Townsville, 4811 Queensland, Australia Received 1 February 2007; received in revised form 31 May 2007; accepted 4 June 2007 Abstract Seagrasses, marine flowering plants, are widely distributed along temperate and tropical coastlines of the world. Seagrasses have key ecological roles in coastal ecosystems and can form extensive meadows supporting high biodiversity. The global species diversity of seagrasses is low (b60 species), but species can have ranges that extend for thousands of kilometers of coastline. Seagrass bioregions are defined here, based on species assemblages, species distributional ranges, and tropical and temperate influences. Six global bioregions are presented: four temperate and two tropical. The temperate bioregions include the Temperate North Atlantic, the Temperate North Pacific, the Mediterranean, and the Temperate Southern Oceans. The Temperate North Atlantic has low seagrass diversity, the major species being Zostera marina, typically occurring in estuaries and lagoons. The Temperate North Pacific has high seagrass diversity with Zostera spp. in estuaries and lagoons as well as Phyllospadix spp. in the surf zone. The Mediterranean region has clear water with vast meadows of moderate diversity of both temperate and tropical seagrasses, dominated by deep-growing Posidonia oceanica. -

Cavitation Resistance Among 26 Chaparral Species of Southern California
Ecological Monographs, 77(1), 2007, pp. 99–115 Ó 2007 by the Ecological Society of America CAVITATION RESISTANCE AMONG 26 CHAPARRAL SPECIES OF SOUTHERN CALIFORNIA 1,4 2 1 3 ANNA L. JACOBSEN, R. BRANDON PRATT, FRANK W. EWERS, AND STEPHEN D. DAVIS 1Department of Plant Biology, Michigan State University, East Lansing, Michigan 48824-1312 USA 2Department of Biology, California State University, Bakersfield, 14 SCI, 9001 Stockdale Highway, Bakersfield, California 93311 USA 3Natural Science Division, Pepperdine University, 24255 Pacific Coast Highway, Malibu, California 90263 USA Abstract. Resistance to xylem cavitation depends on the size of xylem pit membrane pores and the strength of vessels to resist collapse or, in the case of freezing-induced cavitation, conduit diameter. Altering these traits may impact plant biomechanics or water transport efficiency. The evergreen sclerophyllous shrub species, collectively referred to as chaparral, which dominate much of the mediterranean-type climate region of southern California, have been shown to display high cavitation resistance (pressure potential at 50% loss of hydraulic conductivity; P50). We examined xylem functional and structural traits associated with more negative P50 in stems of 26 chaparral species. We correlated raw-trait values, without phylogenetic consideration, to examine current relationships between P50 and these xylem traits. Additionally, correlations were examined using phylogenetic independent contrasts (PICs) to determine whether evolutionary changes in these xylem traits correlate with changes in P50. Co-occurring chaparral species widely differ in their P50 (À0.9 to À11.0 MPa). Species experiencing the most negative seasonal pressure potential (Pmin) had the highest resistance to xylem cavitation (lowest P50). Decreased P50 was associated with increased xylem density, stem mechanical strength (modulus of rupture), and transverse fiber wall area when both raw values and PICs were analyzed. -

Drought-Tolerant and Native Plants for Goleta and Santa Barbara County’S Mediterranean Climate
Drought-Tolerant and Native Plants for Goleta and Santa Barbara County’s Mediterranean Climate Drought tolerant plants for the Santa Barbara and Goleta area. In the 1500's California went through an 80 year drought. During the winter there were blizzards in Central California, the Salinas River froze solid where it flowed into the Monterey Bay. During the summer there was no humidity, no rain, and temperatures in the hundreds for many months. During one year in the 1840's there was no measurable rain in Santa Barbara. (The highest measured rainfall in an hour also was in Southern California, 11 inches in an hour) The same native plants that lived through that are still on the hillsides of California. California native plants that do not normally live in the creeks and ponds are very drought tolerant. The best way to find your plant is to check www.mynativeplants.com and do not water at all. But if you want a simple list of drought tolerant plants that can work for your garden here are some. Adenostoma fasciculatum, Chamise. Adenostoma sparsifolium, Red Shanks Agave deserti, Desert Agave Agave shawii, Coastal Agave Agave utahensis, Century Plant Antirrhinum multiflorum, Multiflowered Snapdragon Arctostaphylos La Panza, Grey Manzanita Arctostaphylos densiflora Sentinel Manzanita Arctostaphylos glandulosa adamsii, Laguna Manzanita. Arctostaphylos crustacea eastwoodiana, Harris Grade manzanita. Arctostaphylos glandulosa zacaensis, San Marcos Manzanita Arctostaphylos glauca, Big Berry Manzanita. Arctostaphylos glauca, Ramona Manzanita Arctostaphylos glauca-glandulosa, Weird Manzanita. 1 | Page Arctostaphylos pungens, Mexican Manzanita Arctostaphylos refugioensis Refugio Manzanita Aristida purpurea, Purple 3-awn Artemisia californica, California Sagebrush Artemisia douglasiana, Mugwort Artemisia ludoviciana, White Sagebrush Asclepias fascicularis, Narrowleaf Milkweed Astragalus trichopodus, Southern California Locoweed Atriplex lentiformis Breweri, Brewers Salt Bush. -

Grow Native Nursery Inventory
Grow Native Nursery Inventory As of Dec 21, 2020 Quantity Scientific Name Common Name Size Available Price Acalypha californica California Copperleaf 4 In 3 $ 6.00 Achillea millefolium 'Sonoma Coast' Sonoma Coast Yarrow 1 Gal 10 $ 10.00 Adenostoma fasciculatum Chamise 1 Gal 10 $ 12.00 Agave sebastiana 'Dwarf Form' Small Form Sebastian's Agave 3 Gal 1 $ 45.00 Agave sebastiana 'Dwarf Form' Small Form Sebastian's Agave 4 In 1 $ 28.00 Alnus rhombifolia White Alder 1 Gal 4 $ 12.00 Aloysia wrightii Oreganillo 1 Gal 3 $ 12.00 Aquilegia formosa Western Columbine 4 In 9 $ 6.00 Arctostaphylos 'Austin Griffiths' Austin Griffiths' Manzanita 1 Gal 11 $ 12.00 Arctostaphylos 'Byrd Hill' Byrd Hill's Manzanita 5 Gal 4 $ 45.00 Arctostaphylos 'Dr. Hurd' Dr. Hurd Manzanita 1 Gal 6 $ 12.00 Arctostaphylos edmundsii 'Bert Johnson' Bert Johnson's Manzanita 1 Gal 15 $ 12.00 Arctostaphylos edmundsii 'Carmel Sur' Carmel Sur Manzanita 4 In 23 $ 6.00 Arctostaphylos glauca Bigberry Manzanita 1 Gal 6 $ 15.00 Arctostaphylos 'Howard Mcminn' Howard McMinn's Manzanita 1 Gal 7 $ 12.00 Arctostaphylos 'Howard Mcminn' Howard McMinn's Manzanita 1 Gal 2 $ 12.00 Arctostaphylos 'Lester Rowntree' Lester Rowntree's Manzanita 1 Gal 11 $ 12.00 Arctostaphylos morroensis Morro Bay Manzanita 1 Gal 3 $ 12.00 Arctostaphylos 'Pacific Mist' Pacific Mist Manzanita 1 Gal 29 $ 12.00 Default Arctostaphylos 'Pt. Reyes' Point Reyes Manzanita Title 10 $ 12.00 Arctostaphylos refugioensis Refugio Manzanita 3 Gal 1 $ 26.00 Arctostaphylos refugioensis Refugio Manzanita 1 Gal 15 $ 12.00 Arctostaphylos -

Arctostaphylos Photos Susan Mcdougall Arctostaphylos Andersonii
Arctostaphylos photos Susan McDougall Arctostaphylos andersonii Santa Cruz Manzanita Arctostaphylos auriculata Mount Diablo Manzanita Arctostaphylos bakeri ssp. bakeri Baker's Manzanita Arctostaphylos bakeri ssp. sublaevis The Cedars Manzanita Arctostaphylos canescens ssp. canescens Hoary Manzanita Arctostaphylos canescens ssp. sonomensis Sonoma Canescent Manzanita Arctostaphylos catalinae Catalina Island Manzanita Arctostaphylos columbiana Columbia Manzanita Arctostaphylos confertiflora Santa Rosa Island Manzanita Arctostaphylos crustacea ssp. crinita Crinite Manzanita Arctostaphylos crustacea ssp. crustacea Brittleleaf Manzanita Arctostaphylos crustacea ssp. rosei Rose's Manzanita Arctostaphylos crustacea ssp. subcordata Santa Cruz Island Manzanita Arctostaphylos cruzensis Arroyo De La Cruz Manzanita Arctostaphylos densiflora Vine Hill Manzanita Arctostaphylos edmundsii Little Sur Manzanita Arctostaphylos franciscana Franciscan Manzanita Arctostaphylos gabilanensis Gabilan Manzanita Arctostaphylos glandulosa ssp. adamsii Adam's Manzanita Arctostaphylos glandulosa ssp. crassifolia Del Mar Manzanita Arctostaphylos glandulosa ssp. cushingiana Cushing's Manzanita Arctostaphylos glandulosa ssp. glandulosa Eastwood Manzanita Arctostaphylos glauca Big berry Manzanita Arctostaphylos hookeri ssp. hearstiorum Hearst's Manzanita Arctostaphylos hookeri ssp. hookeri Hooker's Manzanita Arctostaphylos hooveri Hoover’s Manzanita Arctostaphylos glandulosa ssp. howellii Howell's Manzanita Arctostaphylos insularis Island Manzanita Arctostaphylos luciana -

Strasbourg, 22 May 2002
Strasbourg, 21 October 2015 T-PVS/Inf (2015) 18 [Inf18e_2015.docx] CONVENTION ON THE CONSERVATION OF EUROPEAN WILDLIFE AND NATURAL HABITATS Standing Committee 35th meeting Strasbourg, 1-4 December 2015 GROUP OF EXPERTS ON THE CONSERVATION OF AMPHIBIANS AND REPTILES 1-2 July 2015 Bern, Switzerland - NATIONAL REPORTS - Compilation prepared by the Directorate of Democratic Governance / The reports are being circulated in the form and the languages in which they were received by the Secretariat. This document will not be distributed at the meeting. Please bring this copy. Ce document ne sera plus distribué en réunion. Prière de vous munir de cet exemplaire. T-PVS/Inf (2015) 18 - 2 – CONTENTS / SOMMAIRE __________ 1. Armenia / Arménie 2. Austria / Autriche 3. Belgium / Belgique 4. Croatia / Croatie 5. Estonia / Estonie 6. France / France 7. Italy /Italie 8. Latvia / Lettonie 9. Liechtenstein / Liechtenstein 10. Malta / Malte 11. Monaco / Monaco 12. The Netherlands / Pays-Bas 13. Poland / Pologne 14. Slovak Republic /République slovaque 15. “the former Yugoslav Republic of Macedonia” / L’« ex-République yougoslave de Macédoine » 16. Ukraine - 3 - T-PVS/Inf (2015) 18 ARMENIA / ARMENIE NATIONAL REPORT OF REPUBLIC OF ARMENIA ON NATIONAL ACTIVITIES AND INITIATIVES ON THE CONSERVATION OF AMPHIBIANS AND REPTILES GENERAL INFORMATION ON THE COUNTRY AND ITS BIOLOGICAL DIVERSITY Armenia is a small landlocked mountainous country located in the Southern Caucasus. Forty four percent of the territory of Armenia is a high mountainous area not suitable for inhabitation. The degree of land use is strongly unproportional. The zones under intensive development make 18.2% of the territory of Armenia with concentration of 87.7% of total population. -

Bioregionalism, Community and Environmental Ethics: an Approach to Geographical Borderlines
BIOREGIONALISM, COMMUNITY AND ENVIRONMENTAL ETHICS: AN APPROACH TO GEOGRAPHICAL BORDERLINES Dominique Waissbluth* FTER SOME INTRODUCTORY REMARKS IN SECTION 1 ON THE BIORE- GIONALIST PROJECT, section 2 advances an outline of bioregionalism in view of its implementation in communities. Section 3 examines the concepts of community and locality, assessing their respective relevan- Ace for bioregionalism. In turn, section 4 addresses concerns regarding the aims and scope of bioregionalism. In particular, I intend to demonstrate that the bioregionalist move needs yet further development in order to thoroughly represent a live alternati- ve in the environmental ethical debate on sustainability and borderline. Keywords: bioregionalism; environmental ethics; community; geographical borderlines. BIOREGIONALISMO, COMUNIDAD Y ÉTICA MEDIOAMBIENTAL: UNA APROXIMACIÓN A LOS LÍMITES GEOGRÁFICOS Luego de algunas observaciones introductorias sobre el bioregionalismo en la sec- ción 1, describo las líneas generales de la doctrina bioregionalista en vistas de su implementación en comunidades en la sección 2. La sección 3 examina los con- ceptos de comunidad y localidad, evaluando la relevancia de ellos para el proyecto bioregionalista. La sección 4 aborda los objetivos y los límites del bioregionalismo. * Centre for Applied Philosophy and Public Ethics School of Historical and Philosophical Studies University of Melbourne. Correo electrónico: [email protected] BIOREGIONALISM, COMMUNITY AND ENVIRONMENTAL ETHICS ● DOMINIQUE WAISSBLUTH 13 En particular, intento demostrar que el bioregionalismo requiere de desarrollos y re- finamientos ulteriores para representar una alternativa legitima en el debate en ética medioambiental sobre sustentabilidad y límites. Palabras clave: bioregionalismo; ética medioambiental; comunidad; límites geográficos 1. INTRODUCTORY REMARKS A bioregional project represents an interesting option within the debate on borderli- ne issues, since it deals with a new conception of a more sustainable reorganization of land. -

Arctostaphylos: the Winter Wonder by Lili Singer, Special Projects Coordinator
WINTER 2010 the Poppy Print Quarterly Newsletter of the Theodore Payne Foundation Arctostaphylos: The Winter Wonder by Lili Singer, Special Projects Coordinator f all the native plants in California, few are as glass or shaggy and ever-peeling. (Gardeners, take note: smooth- beloved or as essential as Arctostaphylos, also known bark species slough off old “skins” every year in late spring or as manzanita. This wild Californian is admired by summer, at the end of the growing season.) gardeners for its twisted boughs, elegant bark, dainty Arctostaphylos species fall into two major groups: plants that flowers and handsome foliage. Deep Arctostaphylos roots form a basal burl and stump-sprout after a fire, and those that do prevent erosion and stabilize slopes. Nectar-rich insect-laden not form a burl and die in the wake of fire. manzanita blossoms—borne late fall into spring—are a primary food source for resident hummingbirds and their fast-growing Small, urn-shaped honey-scented blossoms are borne in branch- young. Various wildlife feast on the tasty fruit. end clusters. Bees and hummers thrive on their contents. The Wintershiny, round red fruit or manzanita—Spanish for “little apple”— The genus Arctostaphylos belongs to the Ericaceae (heath O are savored by coyotes, foxes, bears, other mammals and quail. family) and is diverse, with species from chaparral, coastal and (The botanical name Arctostaphylos is derived from Greek words mountain environments. for bear and grape.) Humans use manzanita fruit for beverages, Though all “arctos” are evergreen with thick leathery foliage, jellies and ground meal, and both fruit and foliage have plant habits range from large and upright to low and spreading. -

The Threatened and Near-Threatened Birds of Northern Ussuriland, South-East Russia, and the Role of the Bikin River Basin in Their Conservation KONSTANTIN E
Bird Conservation International (1998) 8:141-171. © BirdLife International 1998 The threatened and near-threatened birds of northern Ussuriland, south-east Russia, and the role of the Bikin River basin in their conservation KONSTANTIN E. MIKHAILOV and YURY B. SHIBNEV Summary Fieldwork on the distribution, habitat preferences and status of birds was conducted in the Bikin River basin, northern Ussuriland, south-east Russia, during May-July 1992,1993, 1995,1996 and 1997. The results of this survey combined with data collected during 1960- 1990, show the area to be of high conservation priority and one of the most important for the conservation of Blakiston's Fish Owl Ketupa blakistoni, Chinese Merganser Mergus squamatus, Mandarin Duck Aix galericulata and Hooded Crane Grus monacha. This paper reports on all of the 13 threatened and near-threatened breeding species of northern Ussuriland, with special emphasis on their occurrence and status in the Bikin area. Three more species, included in the Red Data Book of Russia, are also briefly discussed. Maps show the distribution of the breeding sites of the species discussed. The establishment of a nature reserve in the lower Bikin area is suggested as the only way to conserve the virgin Manchurian-type habitats (wetlands and forests), and all 10 species of special conservation concern. Monitoring of the local populations of Blakiston's Fish Owl, Chinese Merganser and Mandarin Duck in the middle Bikin is required. Introduction No other geographical region of Russia has as rich a biodiversity as Ussuriland which includes the territory of Primorski Administrative Region and the most southern part of Khabarovsk Administrative Region. -

Gulf Plains Bioregion
Gulf Plains bioregion Description Figure 2 Monitoring data coverage Area: 121 100 km2 The Gulf Plains bioregion is characterised by extensive NT alluvial plains and coastal areas. The tropical savanna vegetation comprises mainly eucalypt and tea-tree open woodlands. The regional economy is mainly based on cattle grazing with some prawn fishing, QLD mining and tourism. Tenure is pastoral lease, Aboriginal land and nature reserves. Major population centres are Burketown, Normanton, Karumba and Doomadgee Aboriginal Community. bioregion and sub-IBRA boundary RMDC AussieGRASS data Location The Gulf Plains bioregion is located in the Gulf of Data sourcesMoni availabletoring data coverage Carpentaria in northern Queensland, with a small Data sources include: area (0.7% of bioregion area) in the Northern Territory (NT; see Figures 1 and 2). n Rapid Mobile Data Collection (RMDC) supported by AussieGRASS simulation (of pasture growth Figure 1 Location of the Gulf Plains and utilisation) and remote sensing (Multiple bioregion Regression Bare Ground Index, version bi1); these provide moderate reliability for reporting change (RMDC — road traverses and visual estimates; AussieGRASS — entire rangelands, simulated results with some ground validation) n domestic stocking density, which provides moderate reliability n fire extent, intensity and frequency, which provides high reliability n dust n distance from water n distribution and relative abundance of invasive animals and weeds Location of Gulf Plains bioregion n land use n land values. Gulf Plains bioregion 1 Climate Sustainable management The Gulf Plains bioregion has a monsoonal climate Critical stock forage with a winter dry season and a summer wet season. Spatially averaged median (1890–2005) rainfall is AussieGRASS, levels of simulated pasture 737 mm (April to March rainfall year; see Figure 3).