Occurrence and Diet of Killer Whales in Northern Norway: Seasonal Patterns Relative to the Distribution and Abundance of Norwegian Spring-Spawning Herring
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Marine Envenomations
Environmental Marine envenomations Ingrid Berling Geoffrey Isbister Background The majority of marine envenomings are minor and do Marine stings are common but most are minor and do not not require medical intervention. Jellyfish stings are a require medical intervention. Severe and systemic marine frequent reason for presentation to first aid and primary envenoming is uncommon, but includes box jellyfish stings, healthcare providers. A knowledge of the variety of stings Irukandji syndrome, major stingray trauma and blue-ringed and envenoming syndromes that occur in Australia, octopus envenoming. Almost all marine injuries are caused including those that are clinically significant, and available by jellyfish stings, and penetrating injuries from spiny fish, treatments, is necessary for practitioners, particularly those stingrays or sea urchins. working in coastal regions. Objective This article describes the presentation and management Marine envenoming can be considered in two broad categories: of marine envenomations and injuries that may occur in jellyfish stings and penetrating venomous marine injuries. Jellyfish Australia. stings range from the life-threatening major box jellyfish (Chironex Discussion fleckeri) to painful, but generally benign, bluebottle stings common First aid for jellyfish includes tentacle removal, application to most southeastern Australian beaches (Figure 1). Penetrating of vinegar for box jellyfish, and hot water immersion (45°C venomous marine injuries often occur when handling fish, but can for 20 min) for bluebottle jellyfish stings. Basic life support occur to anyone involved in water activities, fresh water or marine. is essential for severe marine envenomings that result in They are typically more painful than just the trauma of the wound, and cardiac collapse or paralysis. -

Traveler Information
Traveler Information QUICK LINKS Marine Hazards—TRAVELER INFORMATION • Introduction • Risk • Hazards of the Beach • Animals that Bite or Wound • Animals that Envenomate • Animals that are Poisonous to Eat • General Prevention Strategies Traveler Information MARINE HAZARDS INTRODUCTION Coastal waters around the world can be dangerous. Swimming, diving, snorkeling, wading, fishing, and beachcombing can pose hazards for the unwary marine visitor. The seas contain animals and plants that can bite, wound, or deliver venom or toxin with fangs, barbs, spines, or stinging cells. Injuries from stony coral and sea urchins and stings from jellyfish, fire coral, and sea anemones are common. Drowning can be caused by tides, strong currents, or rip tides; shark attacks; envenomation (e.g., box jellyfish, cone snails, blue-ringed octopus); or overconsumption of alcohol. Eating some types of potentially toxic fish and seafood may increase risk for seafood poisoning. RISK Risk depends on the type and location of activity, as well as the time of year, winds, currents, water temperature, and the prevalence of dangerous marine animals nearby. In general, tropical seas (especially the western Pacific Ocean) are more dangerous than temperate seas for the risk of injury and envenomation, which are common among seaside vacationers, snorkelers, swimmers, and scuba divers. Jellyfish stings are most common in warm oceans during the warmer months. The reef and the sandy sea bottom conceal many creatures with poisonous spines. The highly dangerous blue-ringed octopus and cone shells are found in rocky pools along the shore. Sea anemones and sea urchins are widely dispersed. Sea snakes are highly venomous but rarely bite. -

The Polyp and the Medusa Life on the Move
The Polyp and the Medusa Life on the Move Millions of years ago, unlikely pioneers sparked a revolution. Cnidarians set animal life in motion. So much of what we take for granted today began with Cnidarians. FROM SHAPE OF LIFE The Polyp and the Medusa Life on the Move Take a moment to follow these instructions: Raise your right hand in front of your eyes. Make a fist. Make the peace sign with your first and second fingers. Make a fist again. Open your hand. Read the next paragraph. What you just did was exhibit a trait we associate with all animals, a trait called, quite simply, movement. And not only did you just move your hand, but you moved it after passing the idea of movement through your brain and nerve cells to command the muscles in your hand to obey. To do this, your body needs muscles to move and nerves to transmit and coordinate movement, whether voluntary or involuntary. The bit of business involved in making fists and peace signs is pretty complex behavior, but it pales by comparison with the suites of thought and movement associated with throwing a curve ball, walking, swimming, dancing, breathing, landing an airplane, running down prey, or fleeing a predator. But whether by thought or instinct, you and all animals except sponges have the ability to move and to carry out complex sequences of movement called behavior. In fact, movement is such a basic part of being an animal that we tend to define animalness as having the ability to move and behave. -
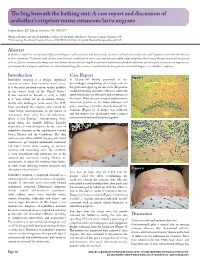
The Bug Beneath the Bathing Suit: a Case Report and Discussion of Seabather’S Eruption Versus Cutaneous Larva Migrans
The bug beneath the bathing suit: A case report and discussion of seabather’s eruption versus cutaneous larva migrans Andrew Jensen, BS,* Marcus Goodman, DO, FAOCD** *Medical Student, 4th year, Philadelphia College of Osteopathic Medicine - Georgia Campus, Suwanee, GA **Dermatology Residency Program Director, PCOM/North Fulton Hospital Medical Campus, Roswell, GA Abstract Seabather’s eruption is an important differential diagnosis when a patient who has recently swum in a subtropical ocean presents with a pruritic rash in the distribution of their swimwear. Treatment with systemic corticosteroids is indicated in severe cases and can successfully reduce symptoms. Oral steroid therapy in general has proven to be an effective treatment for many acute and chronic diseases but has long been associated with increased risk for infections. In this report, we present an atypical case of cutaneous larva migrans and discuss its clinical unmasking after systemic steroid treatment was given for an initial diagnosis of seabather’s eruption. Introduction Case Report Figure 2 Seabather’s eruption is a benign, superficial A 52-year-old female presented to her reaction to toxins from marine-animal larvae. dermatologist complaining of an itchy rash on It is the most common marine-related problem her groin and upper leg for one week. The patient in the waters south of the United States.1 stated she recently traveled to Mexico, where she It was reported in Florida as early as 1903 spent several days on the beach and swimming in as a “rash which set up an intense itching” the ocean. Physical exam revealed erythematous, shortly after bathing in ocean water.2 In 1949, edematous papules on her lower abdomen and Sams postulated the eruption was caused by groin, assuming a location directly beneath her “some living, microorganism, in the nature of swimsuit (Figure 1). -

Marine Mammals and Sea Turtles of the Mediterranean and Black Seas
Marine mammals and sea turtles of the Mediterranean and Black Seas MEDITERRANEAN AND BLACK SEA BASINS Main seas, straits and gulfs in the Mediterranean and Black Sea basins, together with locations mentioned in the text for the distribution of marine mammals and sea turtles Ukraine Russia SEA OF AZOV Kerch Strait Crimea Romania Georgia Slovenia France Croatia BLACK SEA Bosnia & Herzegovina Bulgaria Monaco Bosphorus LIGURIAN SEA Montenegro Strait Pelagos Sanctuary Gulf of Italy Lion ADRIATIC SEA Albania Corsica Drini Bay Spain Dardanelles Strait Greece BALEARIC SEA Turkey Sardinia Algerian- TYRRHENIAN SEA AEGEAN SEA Balearic Islands Provençal IONIAN SEA Syria Basin Strait of Sicily Cyprus Strait of Sicily Gibraltar ALBORAN SEA Hellenic Trench Lebanon Tunisia Malta LEVANTINE SEA Israel Algeria West Morocco Bank Tunisian Plateau/Gulf of SirteMEDITERRANEAN SEA Gaza Strip Jordan Suez Canal Egypt Gulf of Sirte Libya RED SEA Marine mammals and sea turtles of the Mediterranean and Black Seas Compiled by María del Mar Otero and Michela Conigliaro The designation of geographical entities in this book, and the presentation of the material, do not imply the expression of any opinion whatsoever on the part of IUCN concerning the legal status of any country, territory, or area, or of its authorities, or concerning the delimitation of its frontiers or boundaries. The views expressed in this publication do not necessarily reflect those of IUCN. Published by Compiled by María del Mar Otero IUCN Centre for Mediterranean Cooperation, Spain © IUCN, Gland, Switzerland, and Malaga, Spain Michela Conigliaro IUCN Centre for Mediterranean Cooperation, Spain Copyright © 2012 International Union for Conservation of Nature and Natural Resources With the support of Catherine Numa IUCN Centre for Mediterranean Cooperation, Spain Annabelle Cuttelod IUCN Species Programme, United Kingdom Reproduction of this publication for educational or other non-commercial purposes is authorized without prior written permission from the copyright holder provided the sources are fully acknowledged. -

A Guide to Harmful and Toxic Creatures in the Goa of Jordan
Published by the Royal Marine Conservation Society of Jordan. P. O. Box 831051, Abdel Aziz El Thaalbi St., Shmesani 11183. Amman Copyright: © The Royal Marine Conservation Society of Jordan Reproduction of this publication for educational and other non- commercial purposes is authorized without prior written approval from the copyright holder provided the source is fully acknowledged. ISBN: 978-9957-8740-1-8 Deposit Number at the National Library: 2619/6/2016 Citation: Eid, E and Al Tawaha, M. (2016). A Guide to Harmful and Toxic Creature in the Gulf of Aqaba of Jordan. The Royal Marine Conservation Society of Jordan. ISBN: 978-9957-8740-1-8. Pp 84. Material was reviewed by Dr Nidal Al Oran, International Research Center for Water, Environment and Energy\ Al Balqa’ Applied University,and Dr. Omar Attum from Indiana University Southeast at the United State of America. Cover page: Vlad61; Shutterstock Library All photographs used in this publication remain the property of the original copyright holder, and it should not be reproduced or used in other contexts without permission. 1 Content Index of Creatures Described in this Guide ......................................................... 5 Preface ................................................................................................................ 6 Part One: Introduction ......................................................................................... 8 1.1 The Gulf of Aqaba; Jordan ......................................................................... 8 1.2 Aqaba; -

1997 FF13.Pdf
"'\ "',-' ,,- .' ' ... ' ( >J .•; International Council for the ICES C.M. 1997/FF:13 Exploration'of the Sea Fisheries Technology Theme Session ABUNDANCE ESTIMATION OF NORWEGIAN SPRING SPAWNING HERRING WINTERING IN THE VESTFJORD SYSTEM, DECEMBER 1996 by 2 4 Kenneth G. FooteI, Marek Ostrowski , Ingolf R~ttingen3 and Aril Slotte • Institute ofMarine Research P. O. Box 1870 N-5024 Bergen, Norway ABSTRACT The spawning stock ofNorwegian spawning herring has been surveyed when wintering in the Vestfjord, Tysfjord, and Ofotfjord, in northern Norway in December 1996. Tbe basic method is that of echo integration, with compensation for acoustic extinction. Physical capture has been effected by means of the MultisampIer pelagic trawl, which allows remote opening and • closing of each of three codends at arbitrary depths. This has shown essentially thorough mixing of the dominant year classes except in the two fjord arms, where the sampIes contained a larger proportion of older fish (1983 year class). Geostatistics has been employed to describe the spatial structure and to provide an estimate of variance. Data quality control procedures have been performed and are described. 1 e-mail: [email protected] teU: +47 55 23 84 65 fax; +47 55 23 85 32 2 e-mail: [email protected] telf: +47 55 23 6848 fax: +4755 23 83 32 3 e-mail: [email protected]· telf: +47 55238404 fax: +47 55 23 86 87 4 e-mail: [email protected] telf; +4755 23 84 22 fax: +4755 23 86 87 - .. _------------ INTRODUCTION The spawning stock ofNorwegian spring spawning'heinri~'~~'been wintering in the _fjords of northern Norway since 1987. -

The Lillevik Dyke Complex, Narvik: Geochemistry and Tectonic Implications of a Probable Ophiolite Fragment in the Caledonides of the Ofoten Region, North Norway
The Lillevik dyke complex, Narvik: geochemistry and tectonic implications of a probable ophiolite fragment in the Caledonides of the Ofoten region, North Norway ROGNVALD BOYD Boyd, R.: The Lillevik dyke complex, Narvik: geochemistry and tectonic implications of a probable ophiolite fragment in the Caledonides of the Ofoten region, North Norway. Norsk Geologisk Tidsskrift, Vol. 63, pp. 39-54. Oslo 1983, ISSN 0029-196X. The Lillevik dyke complex occurs in an allochthonous unit and shows field relationships indicative of a transition from the mafic cumulate to the sheeted dyke zone in a segment of an ophiolite. Major and trace element chemistry confirm the MORB character of most of the diabases. Certain diabase, gabbro and trondhjemite dykes have REE patterns suggesting a later stage of ocean-island volcanism. The Lillevik complex and equivalent bodies along strike on the eastern limb of the Ofoten synform are a probable source for the mafic facies of the overlying Elvenes Conglomerate. Analogies with other areas suggest that the Lillevik complex was obducted during the Finnmarkian orogeny. R. Boyd, Norges geologiske undersøkelse, Postboks 3006, N-7001 Trondheim, Norway. The topic of this paper is a tectonically bounded gen Groups is marked by a conglomerate hori lens, consisting of gabbro cut by diabase and zon, the Elvenes Conglomerate, which consists gabbroic dykes and by leucocratic veins, which is mainly of matrix-supported cobbles of meta exposed on a shore section within the town of trondhjemite, quartzite and dolomitic marble in Narvik in North Norway. The section Iies in the a matrix of calcareous mica schist (Foslie 1941, upperrnost part of the Narvik Group of Gustav Gustavson 1966); this unit is currently being son (1966, 1972) (Fig. -

NARVIK – Norwegian Eldorado for Wreck-Divers Wrecks of Narvik
NARVIK – Norwegian Eldorado for wreck-divers Wrecks of Narvik Text by Erling Skjold (history and diving) and Frank Bang (diving) Underwater photography by Frank Bang Ship photography by Erling Skjolds, NSA collection Translation by Michael Symes Dieter von Roeder The port of Narvik in north Norway was established around the export of iron-ore from Sweden. This was due to the very good harbour and its ice-free con- ditions. At the outbreak of World War II, Narvik was a strategically important harbour, and during the first few days of the war a very intense battle was fought out here between German, Norwegian and British naval forces. During this fighting several ships were sunk, both warships and civil merchant ships. Narvik harbour was transformed into a great ship ceme- tery, with wrecks sticking up out of the water every- where. Several of the ships were later salvaged, but many wrecks still remained. With its high density of wrecks, Narvik is an eldorado for wreck divers. A diver explores the wreck of the German destroyer Hermann Künne in Trollvika 61 X-RAY MAG : 5 : 2005 EDITORIAL FEATURES TRAVEL NEWS EQUIPMENT BOOKS SCIENCE & ECOLOGY EDUCATION PROFILES PORTFOLIO CLASSIFIED features Narvik Wrecks www.navalhistory.net Narvik harbour Maps outline battles in Narvik and around Norway during World War II Narvik harbour The importance of Narvik as a strate- Attack on April 9th the Eidsvold in just a few seconds. The that it was British gic harbour increased immediately at The German attack was a great surprise German ships could thereafter sail into forces that were the outbreak of World War II. -

K Yanite-Grade Metamorphism in the Evenes and Bogen Groups, Ofoten
Kyanite-grade metamorphism in the Evenes and Bogen Groups, Ofoten, North Norway MARK G. STELTENPOHL & JOHN M. BARTLEY Steltenpohl, M. G. & Bartley, J. M. : Kyanite-grade metamorphism in the Evenes and Bogen Groups, Ofoten, North Norway. Norsk Geologisk Tidsskrift, Vol. 64, pp. 21-26. Oslo 1984. ISSN 0029-196X. Directly north of Ofotfjorden in northern Norway, pelitic schists within the Evenes and Bogen Groups contain the mineral assemblage garnet + biotite ± kyanite ± staurolite + white mica + quartz ± plagio clase. This assemblage implies metamorphic P-T minima of- 540"C and - 4. 8 kb. The rocks are thus at a higher grade than suggested by previous reports, which placed them in the greenschist facies. This indicates that several metamorphic allochthons in Ofoten, including rocks of the Narvik, Evenes, Bogen, and Niingen Groups, are all at kyanite grade, supporting recent interpretations which on structural grounds concluded that the metamorphic peak outlasted stacking of these allochthons. A proposed correlation of the Evenes Group with the Middle Ordovician-Lower Silurian Balsfjord Supergroup implies that this stacking and associated kyanite-grade metamorphism are post-early Silurian and are related to the Scandian phase of the Caledonian orogeny. M. G. Steltenpohl & J. M. Bartley, Department of Geology, University of North Carolina, Chapel Hill, North Carolina 27514, USA. Our mapping, structural analysis, and petro ultramafic rocks of the Narvik Group, which are graphic studies in Ofoten have concentrated on presently at kyanite grade (Foslie 1941, 1949, the structural and metamorphic development of Gustavson 1966, 1972, Hodges 1982a, Tull et al. the Caledonian nappe stack. Figure l shows a in press). -

Our Summer Newsletter Has Gone to the Dogs!
Summer 2013 Newsletter Our summer newsletter has gone to the dogs! In this issue… Pet-Friendly Rentals | Tips for taking your dog to the beach | Beach essentials equipment list | Area vets and boarding | OIB doggy photos | What’s new? and more. There are huge benefits when you travel with your pet. No boarding fees is just one, but there's also no anxiety about how your pet is coping while you're away, less separation stress for your pet, more spacious accommodations that will be just like they're used to at home, and a fun-filled vacation that includes your pet being included with the family. Whether you're a family taking a summer vacation or snowbirds coming south for the winter, you'll be comfortable knowing your pet is welcome here too and that we have a variety of pet friendly rentals to choose from. OCEANFRONT HOUSES 358 East First Street…4 bedroom…$2195 434 East Second St….3 bedroom…$2195 SECOND ROW HOUSES 93 East First Street…..3 bedroom….$965 119 East First Street….4 bedroom…$915 345 East First Street…6 bedroom…$4015 THIRD ROW HOUSES 78 East Second Street..4 bedroom…$995 MID-ISLAND HOUSES 2 Driftwood Street…...2 bedroom…$995 CANAL HOUSES 1 Concord Street……4 bedroom…$1145 41 Concord Street…...4 bedroom…$1315 69 Fairmont Street……4 bedroom…$995 44 Leland Street….…5 bedroom…$1460 26 Wilmington St……6 bedroom…$1095 CONDOS Ocean Cove 105……..1 bedroom…$995 Ocean Cove 219……...1 bedroom…$995 Oceanside West F-1…..3 bedroom…$975 VIOLET, photo courtesy of Julie 1. -

Geochemical Investigations of Northern Norwegian Fjord Sediments – Sources and Spatial Variability of Marine and Terrigenous Components
Faculty of Science and Technology Department of Geology Geochemical investigations of northern Norwegian fjord sediments – Sources and spatial variability of marine and terrigenous components — Lea Philine Canzler Master thesis in Marine Geology and Geophysics (GEO-3900) May 2016 Abstract Fjords represent unique coastal marine environments with high sedimentation rates. They can be used for intensive investigations focusing on geochemical, mineralogical, biological, and sedimentological processes. In this study I investigate 42 surface sediment samples from the outer and inner parts of the Vestfjord, Ofotfjord, Tysfjord and tributary fjord arms in northern Norway. Samples were analysed for their bulk elemental composition, grain size distribution, and total carbon and nitrogen contents (Ctot, Ntot). Furthermore, total organic and inorganic carbon and nitrogen concentrations (Corg, Cinorg, Norg and Ninorg) as well as the carbon and nitrogen isotopes (δ13C, δ15N) were analysed. The measurements were performed to identify geochemical proxies for the contribution of terrestrial organic matter (TOM) versus marine organic matter (MOM) into the fjords. The results indicate a sedimentary environment characterized by an overall dominant contribution of MOM compared to the relatively small input of terrestrial derived material. The extremely high supply of MOM is presented by 13 Corg/Norg ratios ranging between 6 and 10, stable carbon isotope values (δ Corg) between -24‰ 15 and -21‰, and stable organic nitrogen (δ Norg) signatures that vary between 4.7‰ and 6.9‰. The terrigenous material content is in general extraordinary low, and TOM increases from the outer part of the Vestfjord towards the innermost fjord parts of the Ofotfjord and Tysfjord. A 13 15 correlation between δ Corg and δ Norg and the Corg/Norg ratios indicates high influences of marine derived OM.