A Finding of Oligocene Primates on the European Continent
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

I I I I I I I I I I I I I I I I I I
View metadata, citation and similar papers at core.ac.uk brought to you by CORE I provided by NERC Open Research Archive I BRITISH GEOLOGICAL SURVEY I MARINE REPORT SERIES TECHNICAL REPORT WB/95/35 I I I I I I BGS TECHNICAL REPORT WB/95/35 The Wight 1:250 OOO-scale Solid Geology sheet I (2nd Edition) by I I J Andrews I' I Geographical index Wight, English Channel I Subject index: Solid Geology I Production of report was funded by: Science budget I Bibliographic reference: Andrews, U. 1995. The Wight 1:250 OOO-scale Solid Geology sheet (2nd Edition) I British Geological Survey Technical Report WB/95/35 British Geological Survey Tel: 0131 667 1000 I Marine Geology & Operations Group Fax: 0131 6684140 Murchison House Tlx: 727343 West Mains Road I Edinburgh EH93LA I NERC Copyright 1995 I This report has been generated from a scanned image of the document with any blank pages removed at the scanning stage. Please be aware that the pagination and scales of diagrams or maps in the resulting report may not appear as in the original I I CONTENTS Page I' 1. INTRODUCTION 1 I 2. DATASET 2 'II 2.1 Onshore 2 2.2 Offshore 4 I 3. MAP REVISION 9 I 3.1 Amendments 9 3.2 Additional features 10 I 4. GEOLOGY 13 4.1 Structural history of the Wessex-Channel Basin 13 I 4.2 Stratigraphy 14 I 5. HYDROCARBONS 23 I 6. ACKNOWLEDGEMENTS 24 'I 7. REFERENCES AND SELECTED BIBLIOGRAPHY 25 I FIGURES I Figure 1 Location of the GSI deep-seismic survey used during map production I Permo-Triassic isopach map Figure 2 I I I I I I I I I 1. -

The First Record of a Mammal from the Insect Limestone Is a Left Lower Incisor of the Rodent Isoptychus (NHMUK.PV.M45566) (Fig.3B)
Vertebrate remains from the Insect Limestone (latest Eocene), Isle of Wight, UK Hooker, J. J., Department of Earth Sciences, Natural History Museum, Cromwell Road, London, SW7 5BD, UK (corresponding author) Evans, S. E., Department of Cell and Developmental Biology, University College London, Anatomy Building, Gower Street, London, WC1E 6BT, UK Davis, P. G., c/o J. J. Hooker Running head: Insect Limestone vertebrates 1 Abstract A small fauna of vertebrates is recorded from the Insect Limestone, Bembridge Marls Member, Bouldnor Formation, late Priabonian, latest Eocene, of the Isle of Wight, UK. The taxa represented are teleost fishes, lizards including a scincoid, unidentified birds and the theridomyid rodent Isoptychus. The scincoid represents the youngest record of the group in the UK. Of particular note is the taphonomic interpretation based on the preservation of anatomical parts of land-based tetrapods that would have been most likely transported to the site of deposition by wind, namely bird feathers and pieces of shed lizard skin. These comprise the majority of the specimens and suggest that the dominant transport mechanism was wind. Keywords: Bembridge Marls – bird – feather – fish – lizard – mammal – rodent – Scincoidea – skin – Squamata – taphonomic – Theridomyidae 2 The Insect Limestone is a discrete bed of fine-grained, hard, muddy, freshwater to hypersaline limestone near the base of the Bembridge Marls Member of the Bouldnor Formation (Munt 2014; Ross & Self 2014). Its age is late Priabonian, thus latest Eocene (Hooker et al. 2009). Insect and plant remains are relatively common, whilst vertebrate remains are exceptionally rare and are limited to fragmentary skeletal elements of fish, lizard, bird and mammal, bird feathers and pieces of shed lizard skin. -

Devonian Plant Fossils a Window Into the Past
EPPC 2018 Sponsors Academic Partners PROGRAM & ABSTRACTS ACKNOWLEDGMENTS Scientific Committee: Zhe-kun Zhou Angelica Feurdean Jenny McElwain, Chair Tao Su Walter Finsinger Fraser Mitchell Lutz Kunzmann Graciela Gil Romera Paddy Orr Lisa Boucher Lyudmila Shumilovskikh Geoffrey Clayton Elizabeth Wheeler Walter Finsinger Matthew Parkes Evelyn Kustatscher Eniko Magyari Colin Kelleher Niall W. Paterson Konstantinos Panagiotopoulos Benjamin Bomfleur Benjamin Dietre Convenors: Matthew Pound Fabienne Marret-Davies Marco Vecoli Ulrich Salzmann Havandanda Ombashi Charles Wellman Wolfram M. Kürschner Jiri Kvacek Reed Wicander Heather Pardoe Ruth Stockey Hartmut Jäger Christopher Cleal Dieter Uhl Ellen Stolle Jiri Kvacek Maria Barbacka José Bienvenido Diez Ferrer Borja Cascales-Miñana Hans Kerp Friðgeir Grímsson José B. Diez Patricia Ryberg Christa-Charlotte Hofmann Xin Wang Dimitrios Velitzelos Reinhard Zetter Charilaos Yiotis Peta Hayes Jean Nicolas Haas Joseph D. White Fraser Mitchell Benjamin Dietre Jennifer C. McElwain Jenny McElwain Marie-José Gaillard Paul Kenrick Furong Li Christine Strullu-Derrien Graphic and Website Design: Ralph Fyfe Chris Berry Peter Lang Irina Delusina Margaret E. Collinson Tiiu Koff Andrew C. Scott Linnean Society Award Selection Panel: Elena Severova Barry Lomax Wuu Kuang Soh Carla J. Harper Phillip Jardine Eamon haughey Michael Krings Daniela Festi Amanda Porter Gar Rothwell Keith Bennett Kamila Kwasniewska Cindy V. Looy William Fletcher Claire M. Belcher Alistair Seddon Conference Organization: Jonathan P. Wilson -

Late Eocene and Early Oligocene) of the Hampshire Basin
Cainozoic Research, 4(1-2), pp. 27-39, February 2006 The Neritidae of the Solent Group (Late Eocene and Early Oligocene) of the Hampshire Basin M.F. Symonds The Cottage in the Park, Ashtead Park, Ashtead, Surrey KT21 1LE, United Kingdom Received 1 June 2003; revised version accepted 7 March 2005 Gastropods of the family Neritidae in the Solent Group of the Hampshire Basin, southern England are reviewed and two previously unde- scribed taxa are described. New genus: Pseudodostia. New species: Clithon (Pictoneritina) cranmorensis and Clithon (Vittoclithon) headonensis. Neotypes designated for Neritina planulata Edwards, 1866 and Neritina tristis Forbes, 1856. Amended diagnosis: subgenus Vittoclithon. New combinations: Pseudodostia aperta (J. de C. Sowerby, 1823), Clithon (Pictoneritina) concavus (J. de C. Sowerby, 1823), Clithon (Pictoneritina) planulatus (Edwards, 1866) and Clithon (Pictoneritina) bristowi Wenz, 1929. KEY WORDS: Mollusca, Gastropoda, Neritidae, Palaeogene, Hampshire Basin. Introduction Systematic Palaeontology Family Neritidae Rafinesque, 1815 Although the number of species of Neritidae in the Solent Genus Pseudodostia gen. nov. Group is rather limited, specimens are common at certain horizons and they have received the attention of numerous Type species — Nerita aperta J. de C. Sowerby, 1825. Eo- authors in the past. In particular Curry (1960, 265-270) cene, Headon Hill Formation. dealt in detail with the taxonomy of Theodoxus concavus (J. de C. Sowerby, 1823), Theodoxus planulatus (Edwards, Derivatio nominis — The name reflects the close resem- 1866) and Theodoxus bristowi Wenz, 1929. The purpose of blance between the shell of the type species of this genus this paper is to update Curry’s work and to cover additional and that of Nerita crepidularia Lamarck, 1822, the type species. -

Fossil Ants (Hymenoptera: Formicidae): Ancient Diversity and the Rise of Modern Lineages
Myrmecological News 24 1-30 Vienna, March 2017 Fossil ants (Hymenoptera: Formicidae): ancient diversity and the rise of modern lineages Phillip BARDEN Abstract The ant fossil record is summarized with special reference to the earliest ants, first occurrences of modern lineages, and the utility of paleontological data in reconstructing evolutionary history. During the Cretaceous, from approximately 100 to 78 million years ago, only two species are definitively assignable to extant subfamilies – all putative crown group ants from this period are discussed. Among the earliest ants known are unexpectedly diverse and highly social stem- group lineages, however these stem ants do not persist into the Cenozoic. Following the Cretaceous-Paleogene boun- dary, all well preserved ants are assignable to crown Formicidae; the appearance of crown ants in the fossil record is summarized at the subfamilial and generic level. Generally, the taxonomic composition of Cenozoic ant fossil communi- ties mirrors Recent ecosystems with the "big four" subfamilies Dolichoderinae, Formicinae, Myrmicinae, and Ponerinae comprising most faunal abundance. As reviewed by other authors, ants increase in abundance dramatically from the Eocene through the Miocene. Proximate drivers relating to the "rise of the ants" are discussed, as the majority of this increase is due to a handful of highly dominant species. In addition, instances of congruence and conflict with molecular- based divergence estimates are noted, and distinct "ghost" lineages are interpreted. The ant fossil record is a valuable resource comparable to other groups with extensive fossil species: There are approximately as many described fossil ant species as there are fossil dinosaurs. The incorporation of paleontological data into neontological inquiries can only seek to improve the accuracy and scale of generated hypotheses. -

Distribution of Landslides and Geotechnical Properties Within the Hampshire Basin
Distribution of landslides and geotechnical properties within the Hampshire Basin Kwanjai Yuangdetkla, Andy Gibson & Malcolm Whitworth University of Portsmouth, Portsmouth, UK Claire Foster, David Entwisle & Catherine Pennington British Geological Survey, Nottingham UK ABSTRACT This paper outlines the sedimentary sequences and geotechnical properties of the Hampshire Basin, a basin of filled with 700 m of Palaeogene clays, silts, sands and limestones in southern England. The paper presents results so far of a study to synthesize relevant geological and geotechnical data and relate these to the nature of the landslides in this basin. The study has found that stratigraphic sequences and geotechnical properties vary considerably across the basin owing to basin morphology and depositional environments which are correspond to complex paleogeography and tectonic movements during the Tertiary. Over-consolidated clays with low residual shear strengths are extensive on moderately steep slopes and prone to landsliding, especially on over-steepened coastal sections. Landslides vary from mudflows through mudslides, rotational landslides and minor falls. Landslide characteristics are strongly influenced by lithology but gradient appears to be the controlling factor in many cases. The presences of weak strata (clays, lignite, laminated layers), the pre-existing shear surface, the lithological interface (sand overlying clay) play important roles to locally control the position of the shear surface and the type of movements. At a basin scale, inland landslides are associated with the development of drainage system during and since the Tertiary. 1 INTRODUCTION structures trend east-west or northeast-southwest (Melville and Freshney, 1982). Locally, the complex 2 The Hampshire Basin underlies 3 400km of the mainland reactivation of basement faulting has produced many of Southern England and the northern half of the nearby local structures forming gentle hills and valleys such as Isle of Wight (Figure 1). -

Earth and Environmental Science Transactions of the Royal Society of Edinburgh
Earth and Environmental Science Transactions of the Royal Society of Edinburgh http://journals.cambridge.org/TRE Additional services for Earth and Environmental Science Transactions of the Royal Society of Edinburgh: Email alerts: Click here Subscriptions: Click here Commercial reprints: Click here Terms of use : Click here The wasps, bees and ants (Insecta: Vespida=Hymenoptera) from the Insect Limestone (Late Eocene) of the Isle of Wight, UK Alexander V. Antropov, Sergey A. Belokobylskij, Stephen G. Compton, Gennady M. Dlussky, Andrey I. Khalaim, Victor A. Kolyada, Mikhail A. Kozlov, Ksenia S. Perlieva and Alexandr P. Rasnitsyn Earth and Environmental Science Transactions of the Royal Society of Edinburgh / Volume 104 / Issue 3-4 / May 2014, pp 335 - 446 DOI: 10.1017/S1755691014000103, Published online: 30 May 2014 Link to this article: http://journals.cambridge.org/abstract_S1755691014000103 How to cite this article: Alexander V. Antropov, Sergey A. Belokobylskij, Stephen G. Compton, Gennady M. Dlussky, Andrey I. Khalaim, Victor A. Kolyada, Mikhail A. Kozlov, Ksenia S. Perlieva and Alexandr P. Rasnitsyn (2014). The wasps, bees and ants (Insecta: Vespida=Hymenoptera) from the Insect Limestone (Late Eocene) of the Isle of Wight, UK . Earth and Environmental Science Transactions of the Royal Society of Edinburgh, 104, pp 335-446 doi:10.1017/S1755691014000103 Request Permissions : Click here Downloaded from http://journals.cambridge.org/TRE, IP address: 146.231.3.1 on 31 Jul 2014 Earth and Environmental Science Transactions of the Royal Society of Edinburgh, 104, 335–446, 2014 (for 2013) The wasps, bees and ants (Insecta: Vespida ¼ Hymenoptera) from the Insect Limestone (Late Eocene) of the Isle of Wight, UK Alexander V. -

A Review of the Decapod Crustaceans from the Tertiary of the Isle of Wight, Hampshire, U
Bulletin of the Mizunami Fossil Museum, no. 38 (2012), p. 33–51, 4 pls., 2 figs., 2 tables. 33 © 202, Mizunami Fossil Museum A review of the decapod crustaceans from the Tertiary of the Isle of Wight, Hampshire, U. K, with description of three new species W. J. Quayle* and J. S. H. Collins** * 4, Argyll Street, Ryde, Isle of Wight, Hampsire, England<[email protected]> ** 8, Shaws Cottages, Perry Rise, London, SE23 2QN, and The Natural History Museum, London, SW7 5BD, England Abstract To the known decapod crustaceans of the Tertiary deposits of the Isle of Wight, Hampshire, the Early Eocene species, Basinotopus lamarkii (Desmarest) and Dromilites bucklandi Bell are introduced from the mainland, as are Glyphithyreus wetherelli (Bell), Rhachiosoma bispinosum Woodward, Coeloma (Litoricola) dentate (Woodward), Xanthilites bowerbanki Bell, and Xanthopsis unispinosa (M'Coy), the stratigrahic range of which has been extended to the London Clay on the Island. The development of an anomaly concerning Zanthopsis unispinosa, also present, if not peculiar to the island, is well established. A carapace of Harpactocarcinus sp. from this horizon is the first British record for the genus. Late Eocene species new to the island are Goniocypoda quaylei Crane, Orthakrolophus depressus? (Quayle and Collins) and Typilobus belli Quayle and Collins. Three new genera and species are described from the Late Eocene Headon Hill Formation; a callianassid, Vecticallichirus abditus; a goneplacid, Gonioplacoides minuta, and a hexapodid, Headonipus tuberculosus. New, superior material of the Late Eocene species, Typilobus obscurus Quayle and Collins, from the type locality, Colwell Bay necessitates a new description. A table of all known Tertiary species of decapod crustaceans from the Isle of Wight is appended. -

Monocot Fossils Suitable for Molecular Dating Analyses
bs_bs_banner Botanical Journal of the Linnean Society, 2015, 178, 346–374. With 1 figure INVITED REVIEW Monocot fossils suitable for molecular dating analyses WILLIAM J. D. ILES1,2*, SELENA Y. SMITH3, MARIA A. GANDOLFO4 and SEAN W. GRAHAM1 1Department of Botany, University of British Columbia, 3529-6270 University Blvd, Vancouver, BC, Canada V6T 1Z4 2University and Jepson Herbaria, University of California, Berkeley, 3101 Valley Life Sciences Bldg, Berkeley, CA 94720-3070, USA 3Department of Earth & Environmental Sciences and Museum of Paleontology, University of Michigan, 2534 CC Little Bldg, 1100 North University Ave., Ann Arbor, MI 48109-1005, USA 4LH Bailey Hortorium, Plant Biology Section, School of Integrative Plant Science, Cornell University, 410 Mann Library Bldg, Ithaca, NY 14853, USA Received 6 June 2014; revised 3 October 2014; accepted for publication 7 October 2014 Recent re-examinations and new fossil findings have added significantly to the data available for evaluating the evolutionary history of the monocotyledons. Integrating data from the monocot fossil record with molecular dating techniques has the potential to help us to understand better the timing of important evolutionary events and patterns of diversification and extinction in this major and ancient clade of flowering plants. In general, the oldest well-placed fossils are used to constrain the age of nodes in molecular dating analyses. However, substantial error can be introduced if calibration fossils are not carefully evaluated and selected. Here we propose a set of 34 fossils representing 19 families and eight orders for calibrating the ages of major monocot clades. We selected these fossils because they can be placed in particular clades with confidence and they come from well-dated stratigraphic sequences. -

PALAEONTOLOGICAL ASSOCIATION 47Th Annual Meeting Department
PALAEONTOLOGICAL ASSOCIATION 47th Annual Meeting Department of Geology, University of Leicester December 14-17, 2003 ABSTRACTS of TALKS ABSTRACTS of POSTERS Compiled and edited by Mark Purnell Oral presentations Rediscovery of the Gutterford Burn ‘Eurypterid Bed’ Pentland Hills, Midlothian, Scotland Lyall I. Anderson National Museums of Scotland, Chambers St, Edinburgh, EH1 1JF <[email protected]> A programme of field excavation undertaken by National Museums of Scotland staff, volunteers and other interested parties during early July 2003 had three main aims. Firstly to relocate the exact position of the ‘Eurypterid Bed’, a fossiliferous sediment which in the late 1880’s yielded the world’s most diverse assemblage of Silurian chelicerate arthropods, secondly to characterise the likely sedimentary depositional setting and preservational mechanics of this Fossil Konservat-Lagerstätte, and thirdly to investigate the wider relevance of this fossiliferous unit to the more fully known sequences lying stratigraphically higher in the inlier as detailed by the work of Clarkson et al. (2001). After extensive excavation, the ‘Eurypterid Bed’ lithology was located in situ on the banks of the Gutterford Burn stream section. Detailed sedimentary logging and sampling indicated that volcaniclastic sediments played a major role in the formation of the bed; both discrete ashfall bands and ash-rich sediment were discovered in the metre-thick unit along with monograptids. Overlying the ‘eurypterid bed’ occur sporadic horizons yielding dendroid graptolites and numerous (at least 12) thin, discontinuous bands of decalcified marine limestone, rich in invertebrate remains. The fauna within these bands shows a degree of similarity with that identified in the overlying Deerhope Formation. -
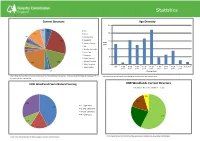
Isle of Wight Forest Plan Statistics and Approval
Statistics Current Structure Age Diversity 250 4% 6% Ash 200 11% Beech 1% Corsican Pine Douglas Fir 4% 23% 150 Lawsons Cypress Area 3% (ha) Oak 2% Western Red Cedar 100 2% Scots Pine Sycamore 50 Sweet Chestnut 16% Western Hemlock 22% Mixed Broadleaf 0 Mixed Conifer 100+ 1-100 81-90 71-80 61-70 51-60 41-50 31-40 21- 30 11-20 0-10 years 4% years years years years years years years years years 2% Planting Years Chart shows the woodland structure broken down into the individual tree species. Only species which make up more than 1% of Chart shows the age structure of the woodland broken down into planting years. the total area are represented. IOW Woodlands Semi Natural Scoring IOW Woodlands Current Structure Broadleaf Conifer Mixed Open 5% 16% 31% 1 = > 80% Native 42% 2 = 50% - 80% Native 3 = 20% - 50% Native 4= < 20% Native 57% 22% 10% 17% Chart shows the current structure of the woodlands separated into generalised habitat types Chart shows the percentage of native species in ancient woodland areas Statistics IOW Woodlands Long Term Structure Average Production Forecast 6000 Broadleaf Open Mixed 5000 4000 26% s e tr e M3000 ic b u C 2000 11% 63% 1000 0 2017-2021 2022-2026 2027-2031 2032-2036 2037-2042 Felling Years Chart shows the long term structure of the woodland separated into generalised habitat types. Time scale is around Chart shows an indicative average timber volume forecast to be produced in 5 year intervals 200 years. Wildfire Risk Assessment A wildfire risk assessment is an evaluation of the likelihood of a wildfire occurring and the severity of damage it might cause if it does occur. -
Brief Report Acta Palaeontologica Polonica 63 (3): 435–440, 2018
Editors' choice Brief report Acta Palaeontologica Polonica 63 (3): 435–440, 2018 A new aneuretine ant from the Paleocene Paskapoo Formation of Canada JOHN S. LAPOLLA and PHILLIP BARDEN While ants were present in the Cretaceous they were rela- present as far back as at least the mid-Cretaceous; ants appear tively rare. This changes dramatically from the Eocene on- to have been a relatively modest component of terrestrial eco- ward when ants rise to the levels of global dominance seen in systems in the Mesozoic (Nel et al. 2004; Perrichot et al. 2008; terrestrial ecosystems today. Their prevalence is well doc- Barden and Grimaldi 2014). Cretaceous ant communities were umented in various Eocene, Oligocene, and Miocene fossil largely comprised of stem-group lineages, which filled sur- deposits, but there is a conspicuous absence of ant-bearing prisingly diverse ecological roles (Barden and Grimaldi 2016), fossil deposits around the Cretaceous–Paleogene bound- but all of which presumably did not survive into the Cenozoic ary. In particular, until this study, there were no definitive following the Cretaceous–Paleogene mass extinctions. While ant-yielding deposits between 78 million and 55 million the ant fossil record is rich, with over 700 described species years ago, a 23-million-year gap. This gap is significant from nearly 70 deposits (Barden 2017), there is a conspicuous because, in addition to spanning a time when ant preva- absence of windows into the Paleocene. Until now, no defini- lence increases by an order of magnitude, the fossil rich tive ant-yielding deposits were known from any ages between deposits from the Eocene onward reveal a dramatically dif- ~78 million-year-old Canadian Medicine Hat amber, the last ferent ant fauna from that observed in the Mesozoic.