Validation of a Method for Age Determination in the Razor Clam Ensis Directus with a Review on Available Data on Growth, Reproduction and Physiology
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Morphometry, Growth and Reproduction of an Atlantic Population of the Razor Clam Ensis Macha (Molina, 1782)*
SCI. MAR., 68 (2): 211-217 SCIENTIA MARINA 2004 Morphometry, growth and reproduction of an Atlantic population of the razor clam Ensis macha (Molina, 1782)* PEDRO J. BARÓN1,2, LUCIANO E. REAL2, NÉSTOR F. CIOCCO1,2 and MARÍA E. RÉ1 1 Centro Nacional Patagónico, CONICET, Boulevard Brown s/n, Puerto Madryn (9120), Chubut, Argentina. E-mail: [email protected] 2 Universidad Nacional de la Patagonia San Juan Bosco, Boulevard Brown 3700, Puerto Madryn (9120), Chubut, Argentina. SUMMARY: Ensis macha is a razor clam distributed throughout the coasts of southern Argentina and Chile. Even though it represents a valuable fishery resource, the exploitation of its Atlantic populations has begun only in recent years. This study provides the first estimates of growth rate, an interpretation of the reproductive cycle on the coast of the northern Argentine Patagonia and an analysis of the species morphometry. Growth was estimated by direct observation of growth rings on the valves by two observers. The reproductive cycle was interpreted by the analysis of temporal change of oocyte size frequency distributions. Parameter estimations for the von Bertalanffy equations respectively obtained by observers 1 -1 and 2 were 154 and 153.7 mm for L∞, 0.25 and 0.20 yr for k, and -0.08 and -0.72 yr for t0. Two spawning peaks were detect- ed: September-November 1999 and May-June 2000. However, mature females were found all year round. An abrupt change in the relationship between shell length and height was detected at 11.2 mm length. Key words: razor clam, morphometric relationships, growth, reproduction, Ensis macha, allometric growth. -

Morphological Variations of the Shell of the Bivalve Lucina Pectinata
I S S N 2 3 47-6 8 9 3 Volume 10 Number2 Journal of Advances in Biology Morphological variations of the shell of the bivalve Lucina pectinata (Gmelin, 1791) Emma MODESTIN PhD of Biogeography, zoology and Ecology University of the French Antilles, UMR AREA DEV ABSTRACT In Martinique, the species Lucina pectinata (Gmelin, 1791) is called "mud clam, white clam or mangrove clam" by bivalve fishermen depending on the harvesting environment. Indeed, the individuals collected have differences as regards the shape and colour of the shell. The hypothesis is that the shape of the shell of L. pectinata (P. pectinatus) shows significant variations from one population to another. This paper intends to verify this hypothesis by means of a simple morphometric study. The comparison of the shape of the shell of individuals from different populations was done based on samples taken at four different sites. The standard measurements (length (L), width or thickness (E - épaisseur) and height (H)) were taken and the morphometric indices (L/H; L/E; E/H) were established. These indices of shape differ significantly among the various populations. This intraspecific polymorphism of the shape of the shell of P. pectinatus could be related to the nature of the sediment (granulometry, density, hardness) and/or the predation. The shells are significantly more elongated in a loose muddy sediment than in a hard muddy sediment or one rich in clay. They are significantly more convex in brackish environments and this is probably due to the presence of more specialised predators or of more muddy sediments. Keywords Lucina pectinata, bivalve, polymorphism of shape of shell, ecology, mangrove swamp, French Antilles. -

Siliqua Patula Class: Bivalvia; Heterodonta Order: Veneroida the Flat Razor Clam Family: Pharidae
Phylum: Mollusca Siliqua patula Class: Bivalvia; Heterodonta Order: Veneroida The flat razor clam Family: Pharidae Taxonomy: The familial designation of this (see Plate 397G, Coan and Valentich-Scott species has changed frequently over time. 2007). Previously in the Solenidae, current intertidal Body: (see Plate 29 Ricketts and Calvin guides include S. patula in the Pharidae (e.g., 1952; Fig 259 Kozloff 1993). Coan and Valentich-Scott 2007). The superfamily Solenacea includes infaunal soft Color: bottom dwelling bivalves and contains the two Interior: (see Fig 5, Pohlo 1963). families: Solenidae and Pharidae (= Exterior: Cultellidae, von Cosel 1993) (Remacha- Byssus: Trivino and Anadon 2006). In 1788, Dixon Gills: described S. patula from specimens collected Shell: The shell in S. patula is thin and with in Alaska (see Range) and Conrad described sharp (i.e., razor-like) edges and a thin profile the same species, under the name Solen (Fig. 4). Thin, long, fragile shell (Ricketts and nuttallii from specimens collected in the Calvin 1952), with gapes at both ends Columbia River in 1838 (Weymouth et al. (Haderlie and Abbott 1980). Shell smooth 1926). These names were later inside and out (Dixon 1789), elongate, rather synonymized, thus known synonyms for cylindrical and the length is about 2.5 times Siliqua patula include Solen nuttallii, the width. Solecurtus nuttallii. Occasionally, researchers Interior: Prominent internal vertical also indicate a subspecific epithet (e.g., rib extending from beak to margin (Haderlie Siliqua siliqua patula) or variations (e.g., and Abbott 1980). Siliqua patula var. nuttallii, based on rib Exterior: Both valves are similar and morphology, see Possible gape at both ends. -

2018 Final LOFF W/ Ref and Detailed Info
Final List of Foreign Fisheries Rationale for Classification ** (Presence of mortality or injury (P/A), Co- Occurrence (C/O), Company (if Source of Marine Mammal Analogous Gear Fishery/Gear Number of aquaculture or Product (for Interactions (by group Marine Mammal (A/G), No RFMO or Legal Target Species or Product Type Vessels processor) processing) Area of Operation or species) Bycatch Estimates Information (N/I)) Protection Measures References Detailed Information Antigua and Barbuda Exempt Fisheries http://www.fao.org/fi/oldsite/FCP/en/ATG/body.htm http://www.fao.org/docrep/006/y5402e/y5402e06.htm,ht tp://www.tradeboss.com/default.cgi/action/viewcompan lobster, rock, spiny, demersal fish ies/searchterm/spiny+lobster/searchtermcondition/1/ , (snappers, groupers, grunts, ftp://ftp.fao.org/fi/DOCUMENT/IPOAS/national/Antigua U.S. LoF Caribbean spiny lobster trap/ pot >197 None documented, surgeonfish), flounder pots, traps 74 Lewis Fishing not applicable Antigua & Barbuda EEZ none documented none documented A/G AndBarbuda/NPOA_IUU.pdf Caribbean mixed species trap/pot are category III http://www.nmfs.noaa.gov/pr/interactions/fisheries/tabl lobster, rock, spiny free diving, loops 19 Lewis Fishing not applicable Antigua & Barbuda EEZ none documented none documented A/G e2/Atlantic_GOM_Caribbean_shellfish.html Queen conch (Strombus gigas), Dive (SCUBA & free molluscs diving) 25 not applicable not applicable Antigua & Barbuda EEZ none documented none documented A/G U.S. trade data Southeastern U.S. Atlantic, Gulf of Mexico, and Caribbean snapper- handline, hook and grouper and other reef fish bottom longline/hook-and-line/ >5,000 snapper line 71 Lewis Fishing not applicable Antigua & Barbuda EEZ none documented none documented N/I, A/G U.S. -

Fish Bulletin 161. California Marine Fish Landings for 1972 and Designated Common Names of Certain Marine Organisms of California
UC San Diego Fish Bulletin Title Fish Bulletin 161. California Marine Fish Landings For 1972 and Designated Common Names of Certain Marine Organisms of California Permalink https://escholarship.org/uc/item/93g734v0 Authors Pinkas, Leo Gates, Doyle E Frey, Herbert W Publication Date 1974 eScholarship.org Powered by the California Digital Library University of California STATE OF CALIFORNIA THE RESOURCES AGENCY OF CALIFORNIA DEPARTMENT OF FISH AND GAME FISH BULLETIN 161 California Marine Fish Landings For 1972 and Designated Common Names of Certain Marine Organisms of California By Leo Pinkas Marine Resources Region and By Doyle E. Gates and Herbert W. Frey > Marine Resources Region 1974 1 Figure 1. Geographical areas used to summarize California Fisheries statistics. 2 3 1. CALIFORNIA MARINE FISH LANDINGS FOR 1972 LEO PINKAS Marine Resources Region 1.1. INTRODUCTION The protection, propagation, and wise utilization of California's living marine resources (established as common property by statute, Section 1600, Fish and Game Code) is dependent upon the welding of biological, environment- al, economic, and sociological factors. Fundamental to each of these factors, as well as the entire management pro- cess, are harvest records. The California Department of Fish and Game began gathering commercial fisheries land- ing data in 1916. Commercial fish catches were first published in 1929 for the years 1926 and 1927. This report, the 32nd in the landing series, is for the calendar year 1972. It summarizes commercial fishing activities in marine as well as fresh waters and includes the catches of the sportfishing partyboat fleet. Preliminary landing data are published annually in the circular series which also enumerates certain fishery products produced from the catch. -

Use of Shell Shape Variation As an Assessment Tool in The
Fisheries Research 186 (2017) 216–222 Contents lists available at ScienceDirect Fisheries Research journal homepage: www.elsevier.com/locate/fishres Use of shell shape variation as an assessment tool in the southernmost razor clam fishery a,b,∗ c,d c Federico Márquez , M. Magdalena Trivellini , Silvina Van der Molen a LARBIM, IBIOMAR − CONICET, Blvd. Brown 2915 (U9120ACD), Puerto Madryn, Argentina b Universidad Nacional de la Patagonia San Juan Bosco, Blvd. Brown 3100, Puerto Madryn (U9120ACD), Chubut, Argentina c IBIOMAR − CONICET, Blvd. Brown 2915 (U9120ACD), Puerto Madryn, Argentina d Universidad Nacional de Córdoba, Av. Vélez Sarsfield 299, Córdoba (×5000JJC), Córdoba, Argentina a r t i c l e i n f o a b s t r a c t Article history: Morphological variation provides a method for phenotypic stock differentiation at inter- and intra- Received 25 April 2016 specific levels. Various methods are used for the assessment of fishery stocks in mollusks; one of them is Received in revised form 25 August 2016 geometric morphometrics. We analyzed morphological variation in razor clams at ten fishing grounds, Accepted 26 August 2016 five from the Argentinean North Patagonian gulfs and five from Chile, and evaluated the occurrence of Handled by A.E. Punt phenotypic stocks between Argentinean and Chilean fisheries, using geometric morphometrics methods. The Argentinean harvesting of Ensis macha is emerging, and represents a way to diversify the shell- Keywords: fisheries in north Patagonia. Nevertheless, fishing and aquaculture play important roles for the Chilean Phenotypic stock Landmarks economy. Various multivariate methods were applied to describe the differences among and between fishing grounds. -

Os Nomes Galegos Dos Moluscos 2020 2ª Ed
Os nomes galegos dos moluscos 2020 2ª ed. Citación recomendada / Recommended citation: A Chave (20202): Os nomes galegos dos moluscos. Xinzo de Limia (Ourense): A Chave. https://www.achave.ga /wp!content/up oads/achave_osnomesga egosdos"mo uscos"2020.pd# Fotografía: caramuxos riscados (Phorcus lineatus ). Autor: David Vilasís. $sta o%ra est& su'eita a unha licenza Creative Commons de uso a%erto( con reco)ecemento da autor*a e sen o%ra derivada nin usos comerciais. +esumo da licenza: https://creativecommons.org/ icences/%,!nc-nd/-.0/deed.g . Licenza comp eta: https://creativecommons.org/ icences/%,!nc-nd/-.0/ ega code. anguages. 1 Notas introdutorias O que cont!n este documento Neste recurso léxico fornécense denominacións para as especies de moluscos galegos (e) ou europeos, e tamén para algunhas das especies exóticas máis coñecidas (xeralmente no ámbito divulgativo, por causa do seu interese científico ou económico, ou por seren moi comúns noutras áreas xeográficas) ! primeira edición d" Os nomes galegos dos moluscos é do ano #$%& Na segunda edición (2$#$), adicionáronse algunhas especies, asignáronse con maior precisión algunhas das denominacións vernáculas galegas, corrixiuse algunha gralla, rema'uetouse o documento e incorporouse o logo da (have. )n total, achéganse nomes galegos para *$+ especies de moluscos A estrutura )n primeiro lugar preséntase unha clasificación taxonómica 'ue considera as clases, ordes, superfamilias e familias de moluscos !'uí apúntanse, de maneira xeral, os nomes dos moluscos 'ue hai en cada familia ! seguir -

Molluscs Gastropods
Group/Genus/Species Family/Common Name Code SHELL FISHES MOLLUSCS GASTROPODS Dentalium Dentaliidae 4500 D . elephantinum Elephant Tusk Shell 4501 D . javanum 4502 D. aprinum 4503 D. tomlini 4504 D. mannarense 450A D. elpis 450B D. formosum Formosan Tusk Shell 450C Haliotis Haliotidae 4505 H. varia Variable Abalone 4506 H. rufescens Red Abalone 4507 H. clathrata Lovely Abalone 4508 H. diversicolor Variously Coloured Abalone 4509 H. asinina Donkey'S Ear Abalone 450G H. planata Planate Abalone 450H H. squamata Scaly Abalone 450J Cellana Nacellidae 4510 C. radiata radiata Rayed Wheel Limpet 4511 C. radiata cylindrica Rayed Wheel Limpet 4512 C. testudinaria Common Turtle Limpet 4513 Diodora Fissurellidae 4515 D. clathrata Key-Hole Limpets 4516 D. lima 4517 D. funiculata Funiculata Limpet 4518 D. singaporensis Singapore Key-Hole Limpet 4519 D. lentiginosa 451A D. ticaonica 451B D. subquadrata 451C Page 1 of 15 Group/Genus/Species Family/Common Name Code D. pileopsoides 451D Trochus Trochidae 4520 T. radiatus Radiate Top 4521 T. pustulosus 4522 T. stellatus Stellate Trochus 4523 T. histrio 4524 T. maculatus Maculated Top 452A T. niloticus Commercial Top 452B Umbonium Trochidae 4525 U. vestiarium Common Button Top 4526 Turbo Turbinidae 4530 T. marmoratus Great Green Turban 4531 T. intercostalis Ribbed Turban Snail 4532 T. brunneus Brown Pacific Turban 4533 T. argyrostomus Silver-Mouth Turban 4534 T. petholatus Cat'S Eye Turban 453A Nerita Neritidae 4535 N. chamaeleon Chameleon Nerite 4536 N. albicilla Ox-Palate Nerite 4537 N. polita Polished Nerite 4538 N. plicata Plicate Nerite 4539 N. undata Waved Nerite 453E Littorina Littorinidae 4540 L. scabra Rough Periwinkle 4541 L. -

Growth Increment Periodicity in the Shell of the Razor Clam Ensis
EGU Journal Logos (RGB) Open Access Open Access Open Access Advances in Annales Nonlinear Processes Geosciences Geophysicae in Geophysics Open Access Open Access Natural Hazards Natural Hazards and Earth System and Earth System Sciences Sciences Discussions Open Access Open Access Atmospheric Atmospheric Chemistry Chemistry and Physics and Physics Discussions Open Access Open Access Atmospheric Atmospheric Measurement Measurement Techniques Techniques Discussions Open Access Biogeosciences, 10, 4741–4750, 2013 Open Access www.biogeosciences.net/10/4741/2013/ Biogeosciences doi:10.5194/bg-10-4741-2013 Biogeosciences Discussions © Author(s) 2013. CC Attribution 3.0 License. Open Access Open Access Climate Climate of the Past of the Past Discussions Growth increment periodicity in the shell of the razor clam Ensis Open Access Open Access directus using stable isotopes as a method to validateEarth age System Earth System Dynamics 1,2 1 1 1 Dynamics2,3 J. F. M. F. Cardoso , G. Nieuwland , R. Witbaard , H. W. van der Veer , and J. P. Machado Discussions 1NIOZ – Royal Netherlands Institute for Sea Research, PO Box 59, 1790 AB Den Burg Texel, the Netherlands 2CIIMAR/CIMAR – Interdisciplinary Centre of Marine and Environmental Research, University of Porto, Rua dos Open Access Open Access Bragas 289, 4050-123 Porto, Portugal Geoscientific Geoscientific 3ICBAS – Instituto de Cienciasˆ Biomedicas´ Abel Salazar, Laboratorio de Fisiologia Aplicada,Instrumentation Universidade do Porto, Instrumentation Rua de Jorge Viterbo Ferreira 228, 4050-313 Porto, Portugal Methods and Methods and Correspondence to: J. F. M. F. Cardoso ([email protected]) Data Systems Data Systems Discussions Open Access Received: 18 February 2013 – Published in Biogeosciences Discuss.: 6 March 2013 Open Access Geoscientific Revised: 31 May 2013 – Accepted: 8 June 2013 – Published: 15 July 2013 Geoscientific Model Development Model Development Discussions Abstract. -

Mussel, Cockle, Variegated Scallop, and Razor Shell) Normally Consumed in Spain, 2005
2237 Journal of Food Protection, Vol. 69, No. 9, 2006, Pages 2237–2240 Copyright ᮊ, International Association for Food Protection Mercury Content in Tinned Molluscs (Mussel, Cockle, Variegated Scallop, and Razor Shell) Normally Consumed in Spain, 2005 ANGEL JOSE´ GUTIE´ RREZ,1* GONZALO LOZANO,1 TOMA´ S GONZA´ LEZ,3 JUAN IGNACIO REGUERA,4 AND ARTURO HARDISSON2 1Department of Animal Biology (Marine Sciences), Faculty of Biology, University of La Laguna, 38206 La Laguna, Tenerife, Canary Islands, Spain; 2Department of Toxicology, Faculties of Pharmacy and Medicine, University of La Laguna, 38071 La Laguna, Tenerife, Canary Islands, Spain; 3Canarian Service of Public Health, Central Laboratory, Santa Cruz de Tenerife, Canary Islands, Spain; and 4Department of Microbiology, Faculty of Food Science and Technology, University of Burgos, Misael Ban˜uelos, Burgos 09001, Spain Downloaded from http://meridian.allenpress.com/jfp/article-pdf/69/9/2237/1678015/0362-028x-69_9_2237.pdf by guest on 26 September 2021 MS 06-053: Received 30 January 2006/Accepted 4 April 2006 ABSTRACT Concentrations of mercury were determined for tinned molluscs (Mollusca, Bivalvia), i.e., mussels (Mytilus spp.), cockles (Cerastoderma edule), variegated scallops (Chlamys varia), and razor shells (Ensis spp.), consumed in Spain. A total of 220 samples were analyzed: 120 mussels, 40 cockles, 24 variegated scallops, and 36 razor shells. Samples were obtained weekly from markets in Santa Cruz de Tenerife (Canary Islands) over a period of 12 months. All observed concentrations of mercury were below the maximum permitted for human consumption (0.5 mg/kg) as defined by European Community Decision 93/ 351/CE. Mercury concentrations were 27.28 Ϯ 12.43 g/kg for mussels, 66.59 Ϯ 23.53 g/kg for cockles, 33.68 Ϯ 15.76 g/kg for variegated scallops, and 21.26 Ϯ 12.24 g/kg for razor shells. -

Damage to Razor Clam (Ensis Siliqua and E
The Glasgow Naturalist (online 2020) Volume 27, Part 3 https://doi.org/10.37208/tgn27303 Herring gull (Larus argentatus) damage to razor clam (Ensis siliqua and E. ensis) shells on the Isle of Cumbrae, Scotland P.G. Moore 32 Marine Parade, Millport, Isle of Cumbrae KA28 0EF E-mail: [email protected] Herring gulls (Larus argentatus ) are an opportunist Fig. 1. Right valves of the razor clams Ensis siliqua (top) and species with a catholic diet (Tinbergen, 1953; Hudson & E. ensis (bottom) from Isle of Cumbrae, Scotland. Note Furness, 1988; Ewins et al., 1994; Moore, 2018). They relative sizes and robustness. (Photo: P.G. Moore) have been reported feeding on razor clams (Ensis leei, was harassed by a great black-backed gull (L. marinus) as "E. directus") in the Dutch Wadden Sea (Cadée, 2000; landing alongside it, which was followed by the herring Enners et al., 2018). Witnessing them feeding on gull flying away with the shell. The herring gulls' initial E. siliqua and E. ensis near my house in Millport, Isle of tendency to open shells at the immediate tide edge Cumbrae, Scotland (NS171547) in April 2020 provided would account for sand grains becoming adherent to the opportunity for comparative observations. flesh fragments found remaining stuck to the inside of the recovered empty shells. Both razor clam species are common in sublittoral sand below the adjacent beach at Kames Bay, Millport (Allen, Most empty shells were orientated with their internal 1962; see Holme, 1951 and Fig . 1 herein for surfaces facing up and the majority (96% of E. -
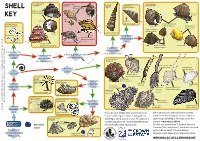
Shell Whelk Dog Whelk Turret It Could Be a Periwinkle Shell (Nucella Lapillus) Shell Spire Shell Thick Top Shell (Osilinus Lineatus) Dark Stripes Key on Body
It could be a type of It could be a type of It could be a It could be a type of topshell whelk Dog whelk turret It could be a periwinkle Shell (Nucella lapillus) shell spire shell Thick top shell (Osilinus lineatus) Dark stripes Key on body Egg Underside capsules Actual size It could be a type of (Hydrobia sp) Common periwinkle spiral worm White ‘Colar’ (Littorina littorea) Flat periwinkle (Littorinasp) Yes Roughly ‘ribbed’ shell. Very high up shore ‘Tooth inside (Turitella communis) opening (Spirorbis sp) Does it have 6 Common whelk No (Buccinum undatum) Yes or more whorls Brown, speckled Netted dog whelk body (twists)? Painted topshell (Nassarius reticulatus) (Calliostoma zizyphinum) No Rough periwinkle Flattened spire Yes Is it long, thin (Littorina saxatilis) Yes Yes and cone shaped Is it permanently No like a unicorn’s horn? attached to Is there a groove or teeth No Is there mother No a surface? in the shell opening? of pearl inside It could be a type of the shell opening? bivalve Yes Yes Common otter-shell (Lutraria lutraria) Bean-like tellin No Is the shell in (Fabulina fabula) Is it 2 parts? spiraled? Common cockle (Cerastoderma edule) It could be a Flat, rounded No sand No Great scallop mason It could be a Is the shell a (Pecten maximus) shell Razor shell worm keel worm Wedge-shaped Is the case dome or (Ensis sp) No Pacific oyster shell made from Yes cone shape? (Crassostrea gigas) Shell can be Peppery furrow shell very large (Scrobicularia plana) sand grains? Elongated and and doesn’t (Lanice conchilega) deep-bodied fully close with large ‘frills’ No (Pomatoceros sp) Yes It could be a type of sea urchin It could be a type of An acorn Native oyster Empty barnacle barnacle Does it have that may be found in estuaries and shores in the UK.