Body Size As a Driver of Scavenging in Theropod Dinosaurs
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Anomalously High Variation in Postnatal Development Is Ancestral for Dinosaurs but Lost in Birds
Anomalously high variation in postnatal development is ancestral for dinosaurs but lost in birds Christopher T. Griffina,1 and Sterling J. Nesbitta aDepartment of Geosciences, Virginia Polytechnic Institute and State University, Blacksburg, VA 24061 Edited by Neil H. Shubin, The University of Chicago, Chicago, IL, and approved November 3, 2016 (received for review August 19, 2016) Compared with all other living reptiles, birds grow extremely fast sequence analysis (OSA) (32) to reconstruct growth sequences of and possess unusually low levels of intraspecific variation during these early dinosaurs, two avian species (Branta canadensis and postnatal development. It is now clear that birds inherited their high Meleagris gallopavo), and a single crocodylian species (Alligator rates of growth from their dinosaurian ancestors, but the origin of mississippiensis), and demonstrate that the earliest dinosaurs the avian condition of low variation during development is poorly developed differently than living archosaurs. constrained. The most well-understood growth trajectories of later Mesozoic theropods (e.g., Tyrannosaurus, Allosaurus)showsimilarly Results low variation to birds, contrasting with higher variation in extant Our OSAs indicate that both C. bauri and M. rhodesiensis pos- crocodylians. Here, we show that deep within Dinosauria, among sessed a high level of intraspecific variation, both in sequence the earliest-diverging dinosaurs, anomalously high intraspecific var- polymorphism and in body size at different levels of morpho- iation is widespread but then is lost in more derived theropods. This logical maturity (Figs. 1 and 2). Analysis of the 27 ontogenetic style of development is ancestral for dinosaurs and their closest characters for C. bauri reconstructed 136 equally parsimonious relatives, and, surprisingly, this level of variation is far higher than developmental sequences (Fig. -

Phylogenetic Analysis of the Allometry of Metabolic Rate and Mitochondrial Basal Proton Leak
Author’s Accepted Manuscript Phylogenetic analysis of the allometry of metabolic rate and mitochondrial basal proton leak Elias T. Polymeropoulos, R. Oelkrug, C.R. White, M. Jastroch www.elsevier.com/locate/jtherbio PII: S0306-4565(16)30294-7 DOI: http://dx.doi.org/10.1016/j.jtherbio.2017.01.013 Reference: TB1878 To appear in: Journal of Thermal Biology Received date: 28 September 2016 Revised date: 24 January 2017 Accepted date: 25 January 2017 Cite this article as: Elias T. Polymeropoulos, R. Oelkrug, C.R. White and M. Jastroch, Phylogenetic analysis of the allometry of metabolic rate and mitochondrial basal proton leak, Journal of Thermal Biology, http://dx.doi.org/10.1016/j.jtherbio.2017.01.013 This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting galley proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain. Phylogenetic analysis of the allometry of metabolic rate and mitochondrial basal proton leak 1 2 3 4,5 Polymeropoulos Elias T. , Oelkrug R. , White C.R. , Jastroch M. 1Menzies Institute for Medical Research, University of Tasmania, 7001 Hobart, Australia, 2Center of Brain, Behaviour and Metabolism, University of Lübeck, 23562 Lübeck, Germany, 3Centre for Geometric Biology, School of Biological Sciences, Monash University, 3800 Melbourne, Australia 4Institute for Diabetes and Obesity, Helmholtz Zentrum, 85764 Munich, Germany, 5 Department of Animal Physiology, Faculty of Biology, Philipps University of Marburg, D-35032 Marburg, Germany Abstract The mitochondrial basal proton leak (MBPL) significantly contributes to high body temperatures (Tb) and basal metabolic rates (BMR) in endotherms. -

Rule Booklet
Dig for fossils, build skeletons, and attract the most visitors to your museum! TM SCAN FOR VIDEO RULES AND MORE! FOSSILCANYON.COM Dinosaurs of North America edimentary rock formations of western North America are famous for the fossilized remains of dinosaurs The rules are simple enough for young players, but and other animals from the Triassic, Jurassic, and serious players can benefit Cretaceous periods of the Mesozoic Era. Your objective from keeping track of the cards that is to dig up fossils, build complete skeletons, and display have appeared, reasoning about them in your museum to attract as many visitors as possible. probabilities and expected returns, and choosing between aggressive Watch your museum’s popularity grow using jigsaw-puzzle and conservative plays. scoring that turns the competition into a race! GAME CONTENTS TM 200,000300,000 160,000 VISITORS VISITORS PER YEAR 140,000 VISITORS PER YEAR 180,000 VISITORS PER YEAR 400,000 VISITORS PER YEAR Dig for fossils, build skeletons, and 340,000 VISITORS PER YEAR RD COLOR ELETONS CA GENUS PERIODDIET SK FOSSIL VISITORSPARTS 360,000 VISITORS PER YEAR PER YEAR attract the most visitors to your museum! VISITORS PER YEAR PER YEAR Tyrannosaurus K C 1 4 500,000 Brachiosaurus J H 1 3 400,000 ON YOUR TURN: TM SCAN FOR VIDEO Triceratops K H 1 3 380,000 RULES AND MORE! Allosaurus J C 2 Dig3 a first360,000 card. If it is a fossil, keep it hidden. FOSSILCANYON.COM Ankylosaurus K H 2 If it3 is an340,000 action card, perform the action. -

A Re-Evaluation of the Enigmatic Dinosauriform Caseosaurus Crosbyensis from the Late Triassic of Texas, USA and Its Implications for Early Dinosaur Evolution
A re-evaluation of the enigmatic dinosauriform Caseosaurus crosbyensis from the Late Triassic of Texas, USA and its implications for early dinosaur evolution MATTHEW G. BARON and MEGAN E. WILLIAMS Baron, M.G. and Williams, M.E. 2018. A re-evaluation of the enigmatic dinosauriform Caseosaurus crosbyensis from the Late Triassic of Texas, USA and its implications for early dinosaur evolution. Acta Palaeontologica Polonica 63 (1): 129–145. The holotype specimen of the Late Triassic dinosauriform Caseosaurus crosbyensis is redescribed and evaluated phylogenetically for the first time, providing new anatomical information and data on the earliest dinosaurs and their evolution within the dinosauromorph lineage. Historically, Caseosaurus crosbyensis has been considered to represent an early saurischian dinosaur, and often a herrerasaur. More recent work on Triassic dinosaurs has cast doubt over its supposed dinosaurian affinities and uncertainty about particular features in the holotype and only known specimen has led to the species being regarded as a dinosauriform of indeterminate position. Here, we present a new diagnosis for Caseosaurus crosbyensis and refer additional material to the taxon—a partial right ilium from Snyder Quarry. Our com- parisons and phylogenetic analyses suggest that Caseosaurus crosbyensis belongs in a clade with herrerasaurs and that this clade is the sister taxon of Dinosauria, rather than positioned within it. This result, along with other recent analyses of early dinosaurs, pulls apart what remains of the “traditional” group of dinosaurs collectively termed saurischians into a polyphyletic assemblage and implies that Dinosauria should be regarded as composed exclusively of Ornithoscelida (Ornithischia + Theropoda) and Sauropodomorpha. In addition, our analysis recovers the enigmatic European taxon Saltopus elginensis among herrerasaurs for the first time. -

A Dinosaur Track from New Jersey at the State Museum in Trenton
New Jersey Geological and Water Survey Information Circular What's in a Rock? A Dinosaur Track from New Jersey at the State Museum in Trenton Introduction a large dinosaur track (fig. 2) on the bottom. Most of the rock is sedimentary, sandstone from the 15,000-foot-thick Passaic A large, red rock in front of the New Jersey State Museum Formation. The bottom part is igneous, lava from the 525-foot- (NJSM) in Trenton (fig. 1) is more than just a rock. It has a thick Orange Mountain Basalt, which overspread the Passaic fascinating geological history. This three-ton slab, was excavated Formation. (The overspreading lava was originally at the top of from a construction site in Woodland Park, Passaic County. It the rock, but the rock is displayed upside down to showcase the was brought to Trenton in 2010 and placed upside down to show dinosaur footprint). The rock is about 200 million years old, from the Triassic footprints Period of geologic time. It formed in a rift valley, the Newark Passaic Formation Basin, when Africa, positioned adjacent to the mid-Atlantic states, began to pull eastward and North America began to pull westward contact to open the Atlantic Ocean. The pulling and stretching caused faults to move and the rift valley to subside along border faults including the Ramapo Fault of northeastern New Jersey, about 8 miles west of Woodland Park. Sediments from erosion of higher Collection site Orange Mountain Basalt top N Figure 1. Rock at the New Jersey State Museum. Photo by W. Kuehne Adhesion ripples DESCRIPTION OF MAP UNITS 0 1 2 mi Orange Mountain Basalt L 32 cm Jo (Lower Jurassic) 0 1 2 km W 25.4 cm contour interval 20 feet ^p Passaic Formation (Upper Triassic) Figure 3. -

Dinosaur Metabolism and the Allometry of Maximum Growth Rate
RESEARCH ARTICLE Dinosaur Metabolism and the Allometry of Maximum Growth Rate Nathan P. Myhrvold* Intellectual Ventures, Bellevue, Washington, United States of America * [email protected] Abstract a11111 The allometry of maximum somatic growth rate has been used in prior studies to classify the metabolic state of both extant vertebrates and dinosaurs. The most recent such studies are reviewed, and their data is reanalyzed. The results of allometric regressions on growth rate are shown to depend on the choice of independent variable; the typical choice used in prior studies introduces a geometric shear transformation that exaggerates the statistical OPEN ACCESS power of the regressions. The maximum growth rates of extant groups are found to have a Citation: Myhrvold NP (2016) Dinosaur great deal of overlap, including between groups with endothermic and ectothermic metabo- Metabolism and the Allometry of Maximum lism. Dinosaur growth rates show similar overlap, matching the rates found for mammals, Growth Rate. PLoS ONE 11(11): e0163205. reptiles and fish. The allometric scaling of growth rate with mass is found to have curvature doi:10.1371/journal.pone.0163205 (on a log-log scale) for many groups, contradicting the prevailing view that growth rate Editor: Anthony Fiorillo, Perot Museum of Nature allometry follows a simple power law. Reanalysis shows that no correlation between growth and Science, UNITED STATES rate and basal metabolic rate (BMR) has been demonstrated. These findings drive a con- Received: January 7, 2015 clusion that growth rate allometry studies to date cannot be used to determine dinosaur Accepted: September 6, 2016 metabolism as has been previously argued. -

Teacher Resource For: Evidence for Mesothermy in Dinosaurs
Teacher Resource for: Evidence for mesothermy in dinosaurs Table of Contents: I. Student Learning Goals II. Connect to Learning Standards III. Summary of the Article for the Teacher IV. Discussion Questions V. Related Multimedia Resources from HHMI|BioInteractive NOTE: To explore mass extinctions, including the end of the dinosaurs, we recommend HHMI's free eBook Mass Extinctions: Lessons from the Past. Student Learning Goals Connections to the nature of science from the article Were dinosaurs slow-metabolizing ectotherms like reptiles or fast-metabolizing endotherms like mammals and birds whose activities were unconstrained by temperature? How can the resting metabolic rate of extinct species be determined using fossils? The importance of this scientific research This research furthers the understanding of how dinosaurs regulated their body temperature in comparison to known ectotherms and endotherms. It determines the resting metabolic rate of dinosaurs using maximum growth rate as measured by bone growth rings. The actual science involved Thermodynamics Paleontology Connect to Learning Standards: The Next Generation Science Standards Science and Engineering Practice 2: Developing and using models Science and Engineering Practice 6: Constructing explanations (for science) and designing solutions (for engineering) The AP Biology Standards Essential knowledge 1.A.4: Biological evolution is supported by scientific evidence from many disciplines, including mathematics. Essential knowledge 2.A.1: All living systems require constant input of free energy. Essential knowledge 2.C.2: Organisms respond to changes in their external environments. Essential knowledge 2.D.2: Homeostatic mechanisms reflect both common ancestry and divergence due to adaptation in different environments. Common Core English Language Arts 11.12.8: Evaluate the hypotheses, data, analyses, and conclusions in a science or technical test, verifying the data when possible and corroborating or challenging conclusions with other sources of information. -

Cranial Anatomy of Allosaurus Jimmadseni, a New Species from the Lower Part of the Morrison Formation (Upper Jurassic) of Western North America
Cranial anatomy of Allosaurus jimmadseni, a new species from the lower part of the Morrison Formation (Upper Jurassic) of Western North America Daniel J. Chure1,2,* and Mark A. Loewen3,4,* 1 Dinosaur National Monument (retired), Jensen, UT, USA 2 Independent Researcher, Jensen, UT, USA 3 Natural History Museum of Utah, University of Utah, Salt Lake City, UT, USA 4 Department of Geology and Geophysics, University of Utah, Salt Lake City, UT, USA * These authors contributed equally to this work. ABSTRACT Allosaurus is one of the best known theropod dinosaurs from the Jurassic and a crucial taxon in phylogenetic analyses. On the basis of an in-depth, firsthand study of the bulk of Allosaurus specimens housed in North American institutions, we describe here a new theropod dinosaur from the Upper Jurassic Morrison Formation of Western North America, Allosaurus jimmadseni sp. nov., based upon a remarkably complete articulated skeleton and skull and a second specimen with an articulated skull and associated skeleton. The present study also assigns several other specimens to this new species, Allosaurus jimmadseni, which is characterized by a number of autapomorphies present on the dermal skull roof and additional characters present in the postcrania. In particular, whereas the ventral margin of the jugal of Allosaurus fragilis has pronounced sigmoidal convexity, the ventral margin is virtually straight in Allosaurus jimmadseni. The paired nasals of Allosaurus jimmadseni possess bilateral, blade-like crests along the lateral margin, forming a pronounced nasolacrimal crest that is absent in Allosaurus fragilis. Submitted 20 July 2018 Accepted 31 August 2019 Subjects Paleontology, Taxonomy Published 24 January 2020 Keywords Allosaurus, Allosaurus jimmadseni, Dinosaur, Theropod, Morrison Formation, Jurassic, Corresponding author Cranial anatomy Mark A. -

Junior Paleontologists Journal – Week 3
Junior Paleontologists Journal – Week 3 Welcome back, Junior Paleontologists! Are you ready to continue your adventures with dinosaurs and other fossils for another week? Dinosaur Detectives: Did you figure out what the tracks of Dilophosaurus looked like? Here is a picture of Dilophosaurus tracks from our exhibit at MNA: Does this look like the tracks you drew or imagined? There are some tracks like these in sandstones near Tuba City, Arizona. Now for Week 3: Our first dinosaur was Coelophysis from the Triassic Period. Our second was Dilophosaurus from the Jurassic Period. This week we will look at a dinosaur from the Cretaceous Period. It is a very different dinosaur, with the very difficult name Nothronychus. Besides its strange name, this dinosaur might be the strangest, weirdest dinosaur in the world! This is my favorite middle-size dinosaur because it is so weird, and because with a field crew I excavated it from southern Utah. This one is a little bigger than Dilophosaurus from last week, and much heavier. We have a complete skeleton on display at the Museum of Northern Arizona. First, let’s have some fun with two very hard words: “Nothronychus” and “therizinosaurs.” This dinosaur’s name is Nothronychus. Say it out loud: NO- throw-NIE-kus. Practice that, say it three times in a row. NO-throw-NIE-kus NO-throw-NIE-kus NO-throw-NIE-kus This dinosaur has a family with other dinosaurs, all of them very weird. The family is called the “therizinosaurs” or THAIR-uh-ZINE-o-saurs. The first part of the name sounds like the TH in “throw.” Now say it three times in a row. -

A Juvenile Coelophysoid Skull from the Early Jurassic of Zimbabwe, and the Synonymy of Coelophysis and Syntarsus
A juvenile coelophysoid skull from the Early Jurassic of Zimbabwe, and the synonymy of Coelophysis and Syntarsus Anthea Bristowe* & Michael A. Raath Bernard Price Institute for Palaeontological Research, School of Geosciences, University of the Witwatersrand, Private Bag 3, WITS, 2050 South Africa Received 23 September 2004. Accepted 5 December 2004 Several authors have drawn attention to the close similarities between the neotheropod dinosaurs Coelophysis and Syntarsus. Recon- struction and analysis of a skull from a juvenile specimen of Syntarsus (collected from the Forest Sandstone Formation of Zimbabwe) show that cranial characters previously used to distinguish these taxa and justify their generic separation (namely the presence of a ‘nasal fenestra’ in Syntarsus and the length of its antorbital fenestra), were based on erroneous reconstructions of disassociated cranial elements. On the basis of this reinterpretation we conclude that Syntarsus is a junior synonym of Coelophysis. Variations are noted in three cranial characters – the length of the maxillary tooth row, the width of the base of the lachrymal and the shape of the antorbital maxillary fossa – that taken together with the chronological and geographical separation of the two taxa justify separation at species level. Keywords: Dinosaurs, Neotheropoda, Coelophysoid, taxonomy, Triassic, Jurassic. INTRODUCTION Following the work of Gauthier (1986), these taxa were Ever since the theropod Syntarsus rhodesiensis was first suggested to belong to a monophyletic clade known as described (Raath 1969), a succession of authors have Ceratosauria. However, more recent works by a number commented on the close morphological similarity be- of authors (Sereno 1997, 1999; Holtz 2000; Wilson et al. tween it and Coelophysis bauri (Raath 1969, 1977; Paul 1988, 2003; Rauhut 2003) have re-evaluated theropod interrela- 1993; Colbert 1989; Rowe 1989; Tykoski 1998; Downs tionships. -
Mesozoic Era Part 1 Sign & Return
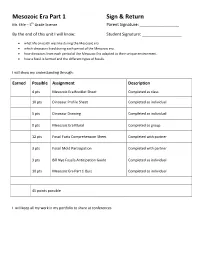
Mesozoic Era Part 1 Sign & Return Mr. Ehle – 5 th Grade Science Parent Signature: ______________________ By the end of this unit I will know: Student Signature: ______________________ • what life on earth was like during the Mesozoic era • which dinosaurs lived during each period of the Mesozoic era. • how dinosaurs from each period of the Mesozoic Era adapted to their unique environment. • how a fossil is formed and the different types of fossils. I will show my understanding through: Earned Possible Assignment Description 4 pts Mesozoic Era Booklet Sheet Completed as class 10 pts Dinosaur Profile Sheet Completed as individual 5 pts Dinosaur Drawing Completed as individual 0 pts Mesozoic Era Mural Completed as group 12 pts Fossil Facts Comprehension Sheet Completed with partner 3 pts Fossil Mold Participation Completed with partner 3 pts Bill Nye Fossils Anticipation Guide Completed as individual 10 pts Mesozoic Era Part 1 Quiz Completed as individual 45 points possible I will keep all my work in my portfolio to share at conferences. Mesozoic Era Part 1 Study Guide Vocabulary Mesozoic Era An era of geologic time that lasted from 245-65 million years ago Triassic Period 245-208 Million Years Ago Jurassic Period 208-135 Million Years Ago Cretaceous Period 135-65 Million Years Ago adapt To adjust to particular conditions extinction The total disappearance of a species so that it no longer exists amber tree sap that became hard fossil bones and other signs of a plant or animal paleontologist scientist who studies fossils trace fossil fossil that shows just a sign of something, like a footprint The Mesozoic Era is so long that it is divided into 3 parts: Triassic, Jurassic and Cretaceous periods. -

A New Theropod Dinosaur from the Early Jurassic of South Africa and Its Implications for the Early Evolution of Theropods
A new theropod dinosaur from the Early Jurassic of South Africa and its implications for the early evolution of theropods Adam M. Yates Bernard Price Institute for Palaeontological Research, School of Geosciences, University of the Witwatersrand, Private Bag 3, WITS 2050, Johannesburg, South Africa E-mail: [email protected] Received 27 June 2005. Accepted 21 September 2005 A new theropod, Dracovenator regenti, from the upper Elliot Formation is described, based upon a fragmentary skull. It can be diagnosed on the basis of a bilobed fossa on the lateral surface of the premaxilla that is connected to the alveolar margin by a narrow channel, the presence of a deep, oblique, lateral notch on the articular and hypertrophied dorsal processes on the articular. Other aspects of its morphology display a mosaic of coelophysoid and advanced theropod characteristics. A cladistic analysis of basal Theropoda, including the new taxon finds that the new taxon is closely related to Dilophosaurus wetherilli and Zupaysaurus rougieri although the clade formed by these three taxa is not robustly supported. It also finds that Coelophysoidea sensu lato is paraphyletic with respect to Ceratosauria + Tetanurae but that this topology is not a significantly better explanation of the data than an inclusive, monophyletic Coelophysoidea. Keywords: Theropoda, Coelophysoidea, Dracovenator, upper Elliot Formation, South Africa. INTRODUCTION 2004). It is now the majority view amongst theropod Prior to Gauthier’s classic (1986) monograph, our under- systematists that Ceratosauria contains Ceratosaurus spp. standing of the interrelationships of theropod dinosaurs and Abelisauroidea and that this clade is more closely could be described as murky at best.